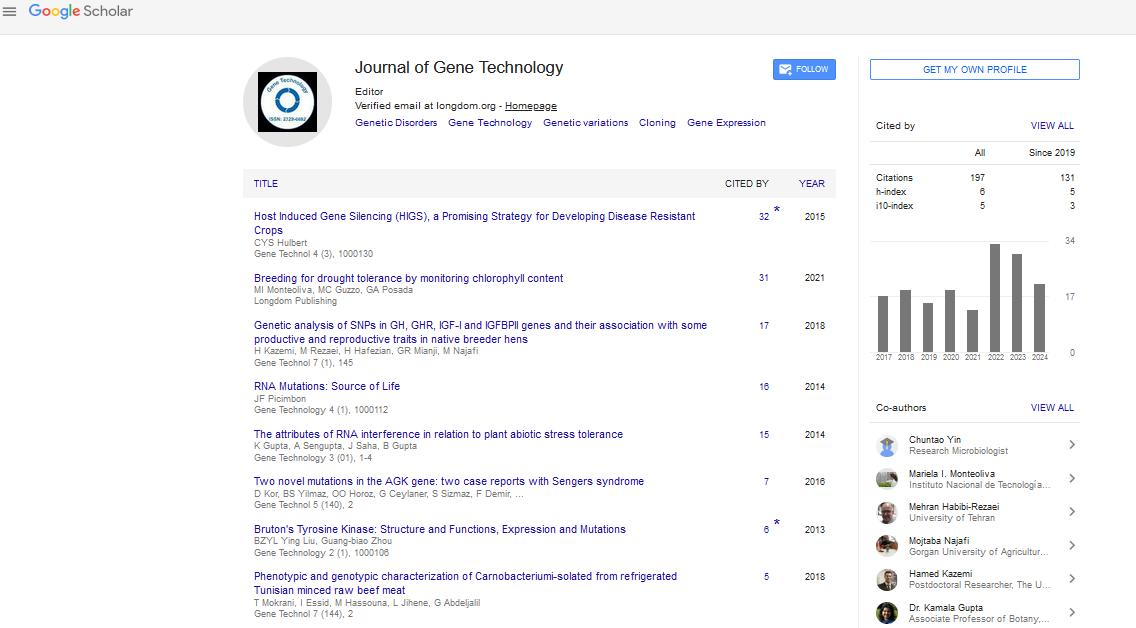

Indexed In
- Academic Keys
- ResearchBible
- CiteFactor
- Access to Global Online Research in Agriculture (AGORA)
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Perspective - (2023) Volume 12, Issue 1
Transcriptome Evolution with Ageing is Shaped by Genome-Wide RNA Polymerase
George Adler*Received: 02-Feb-2023, Manuscript No. RDT-23-20059; Editor assigned: 06-Feb-2023, Pre QC No. RDT-23-20059 (PQ); Reviewed: 20-Feb-2023, QC No. RDT-23-20059; Revised: 27-Feb-2023, Manuscript No. RDT-23-20059 (R); Published: 06-Mar-2023, DOI: 10.35248/2329-6682.23.12.213
Description
Aging is characterised by a progressive loss in molecular, cellular, and physiological function, which results in diminished vitality, age-related illnesses, and increased mortality. Only few things were recognized about the functional status of the basal transcription process in ageing, despite the fact that numerous processes slow down or change as we age. Brains of older rat and fruit fly produce fewer messenger RNAs, and cell-to-cell variation in transcription is increased in various tissues, whereas gene-togene transcriptional coordination is reduced in aging. However, the majority of studies on transcription in ageing focuses on changes in gene expression. Transcriptomics has significantly assisted in the identification of several cellular pathways and processes involved in ageing. Transcriptional factors, changing cell type composition from microRNAs and epigenetic modifications can all contribute to age-related, organ-specific gene expression variations. The majority of the similarities in gene expression between the organs of aged mice could not be attributable to these established regulatory mechanisms, according to a recent transcriptomics meta-analysis.
The accumulation of DNA damage has been proposed as an underlying cause of normal ageing and the aforementioned transcriptional phenotypes, based mostly on parallels to cells exposed to DNA-damaging chemicals or premature ageing DNA repair diseases such as Cockayne syndrome and trichothiodystrophy. Transcription-Coupled Repair (TCR) deficiencies in these diseases result in stopped RNA polymerases on DNA lesions, which suggests that transcription-blocking DNA damage may also play a role in healthy ageing. Although endogenous transcription-blocking DNA lesions build up with normal ageing, it is not yet known if they significantly affect transcription. We used an nascent RNA sequencing approach paired with RNA Polymerase II (RNAPII) Chromatin Immuno Precipitation followed by sequencing (ChIP-seq) and confocal imaging to investigate the basal transcription underpinning gene expression variations in normal wild-type aged mice. By accumulating DNA damage as a general ageing phenotype, we show a severe age-related transcriptional reduction and skewing of transcriptional output, producing age-related transcription alterations generally and especially affecting life span-determining ageing signature pathways.
In order to find a mechanism explaining RNAPII stalling, we then investigated if certain potential characteristics. Nucleotide content across gene bodies, transcriptional error rate, alternative splicing, chromatin accessibility, histone modifications associated with euchromatin, or DNA methylation patterns, which would suggest epigenetic changes are to blame, were not significantly different between genes and other gene categories. These factors do not explain the observed transcriptional reduction since they do not correlate with the level which is predicted when such a component is causally implicated. Given that lengthy genes are more likely to have stochastic lesions, accumulation of transcription-blocking DNA damage is a feasible explanation for gene-length-dependent transcriptional pausing.
It is anticipated that during the strand-specific DNA amplification step in the RNAPII ChIP-seq library protocol, the lesion in the template strand that actually stalls the RNAPII will impair DNA amplification of that strand, in contrast to the undamaged (coding) strand, if a significant portion of elongating RNAPII in ageing is stalled by endogenous transcription-blocking lesions. As seen for UV-induced transcription-blocking lesions, this should result in a strand amplification bias in favour of the coding strand that can be seen by strand-specific ChIP-seq.
First, we established that using our ChIP-seq approach, UVinduced DNA damage causes a coding strand bias that eventually goes away. Significantly, we found in the whole and RNAPIIser2p ChIP-seq datasets an age-related gene-length-dependent coding strand bias in elderly livers. Indicating that polymeraseblocking perturbations are present in the isolated, purified DNA from old livers, regions with a strong coding strand bias showed both unaltered local DNA methylation status or nucleotide content, designating them as damaged DNA. Moreover, the coding strand bias associated with ageing increased towards the ends of genes, particularly in lengthy genes, which was correlated with RNAPII stalling in gene bodies and TCR being more active at the start of genes.
The absence of coding strand bias in the first third of the gene body, which was distinguished by normal or even greater nascent RNA and RNAPII levels, suggests that coding strand bias and transcriptional decrease are correlated. Inferring that promoter silencing is not the cause of reduced IGF1 expression in old liver, DNA damage-induced transcriptional stress is. We estimated the first-to-last exon loss in nascent RNA fold in ageing across all expressed genes and were gene-length-dependent to quantify the effects of transcriptional stress on exons genomewide. This was in line with older people producing less mRNA, offering a possible explanation for the previously noted decline in cellular mRNA levels. This suggests that as we age, transcriptional output declines and gene expression is skewed towards small genes. We looked at which cellular functions and pathways were most vulnerable to transcriptional stress since it decreases and skews transcriptional output. A B cell signature, which denotes age-related B cell infiltration8 and also showed transcriptional stress, was present in the promoter-up regulated gene set.
Normal aging-related transcription-blocking DNA damage results in frequent genome-wide elongating RNAPII stalling, which in turn causes reduced, gene-length-dependent transcriptional output and dysregulation of numerous aging-related processes. We predict that an initial RNAPII stalled on a lesion will block about three more RNAPII complexes causing queuing based on transcription-stalling similarities in UV-treated cells. The underlying processes that cause changes in gene expression with ageing have largely eluded researchers, and they are frequently believed to be the consequence of active regulatory systems such promoter regulation, as well as in DNA repair mutant animal models of premature ageing. However, we believe that passive transcriptional stress caused by DNA damage, in conjunction with gene architecture, i.e. gene length, accounts for a significant portion of these alterations.
Citation: Adler G (2023) Transcriptome Evolution with Ageing is Shaped by Genome-Wide RNA Polymerase. Gene Technol. 12:213.
Copyright: © 2023 Adler G. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.