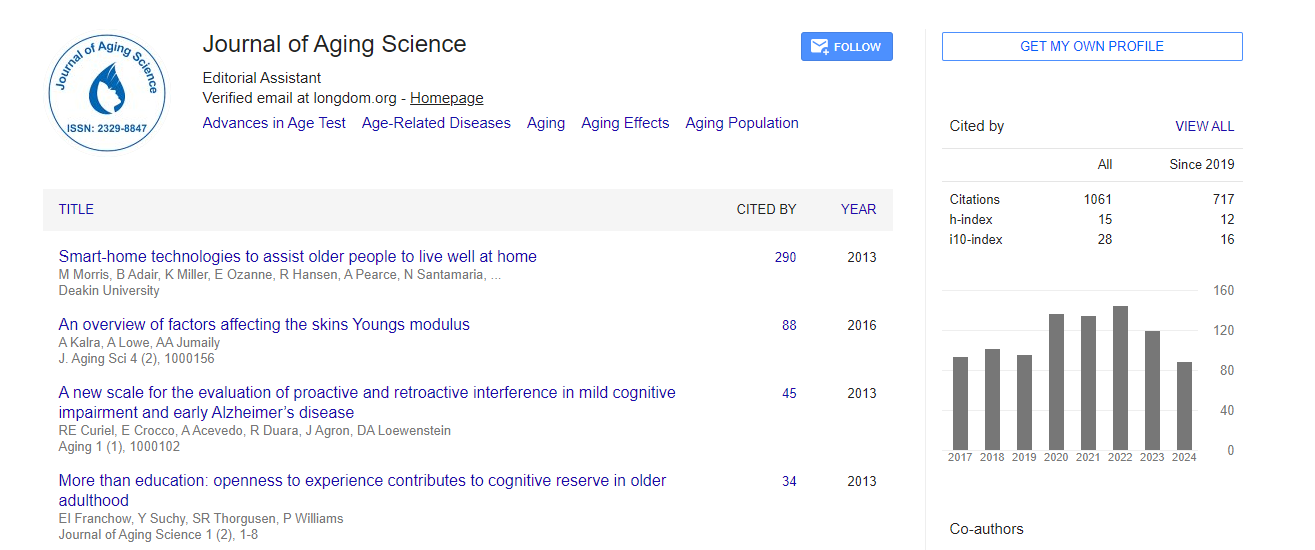

Indexed In
- Open J Gate
- Academic Keys
- JournalTOCs
- ResearchBible
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Research Article - (2022) Volume 10, Issue 3
The Role of Growth Factors in Articular Cartilage Repair: A Systematic Review
Yu Zhang1*, Zishu Cai2, Chengqiang Y1, Youcai Wu1, Yufu Ou1 and Jianxun Wei12Department of Orthopaedics, Guangxi University of Chinese Medicine, Nanning City, Guangxi Zhuang Autonomous Region, China
Received: 27-Apr-2022, Manuscript No. JASC-22-15758; Editor assigned: 29-Apr-2022, Pre QC No. JASC-22-15758(PQ); Reviewed: 16-May-2022, QC No. JASC-22-15758; Revised: 23-May-2022, Manuscript No. JASC-22-15758(R); Published: 30-May-2022, DOI: 10.35248/2329-8847.22.10.274
Abstract
Natural degeneration or trauma can lead to structural and functional damage to articular cartilage. Because cartilage lacks blood supply and innervation, the metabolic activity of its cells is low, and post injury self-repair is difficult. Notably, growth factors have been found to greatly affect articular cartilage repair. English-language studies published from August 2000 to August 2019 were searched in the PubMed and SCI databases. The relevant literature was reviewed, and basic research and clinical applications of growth factors in articular cartilage repair were analysed and summarized. Growth factors significantly promote stem cell proliferation and differentiation and induce their functions. Various growth factors synergistically promote chondrogenic differentiation of stem cells to promote cartilage regeneration and repair articular cartilage damage. Traditional growth factors that promote articular cartilage repair include bone morphogenetic proteins, cartilage-derived morphogenetic proteins, transforming growth factor β, fibroblast growth factors and insulin-like growth factors. Recent studies have shown that kartogenin, platelet-rich plasma, platelet-rich fibrin, mechano-growth factor, etc. can effectively induce chondrogenic differentiation of stem cells and chondrocyte phenotype maintenance; synthetic compounds, e.g., dexamethasone and some inorganic particles, also promote chondrogenic differentiation. Different hydrogel types and stem cells from different sources differentially support chondrogenesis and require different growth factors to induce chondrogenic differentiation. Novel growth factors were found to promote articular cartilage repair. As no in vivo experimental studies have yet addressed dexamethasone and inorganic particles, their reparative effect and safety require further study. The synergism and antagonism between different growth factors and optimal concentration ratios, as well as differences in their in vivo and in vitro roles, also warrant in-depth study.
Keywords
Articular cartilage; Growth factor; Stem cells; Stromal cells
Introduction
After knee joint injury, articular cartilage defects (ACDs) can progress to osteoarthritis (OA), which is very common in China and worldwide. According to relevant statistics from the WHO, 90% of women and 80% of men over the age of 65 years suffer from OA [1,2]. Articular cartilage is an important component in the smooth, painless and functional movement of joints. It is one of the few self-healing tissues in the human body that undergo thinning due to disease or aging, and traumatic injury may lead to local defects [3]. However, articular cartilage is well-hydrated hyaline cartilage. It has neither nerves nor blood vessels, and its low cell density allows only extremely limited self-renewal. Chondrocytes in matrix lacunae obtain necessary nutrients and excrete metabolites mainly through infiltration. The metabolic activity of chondrocytes is low, and their lack of blood supply makes articular cartilage repair very difficult. ACDs often manifest as intractable pain and progressive joint dysfunction, which may eventually lead to degenerative osteoarthritis (OA) [4].In articular cartilage, both acute injury and slow progression of OA cause pain, leading to progressive destruction of the whole joint and resulting in partial or complete loss of joint function [5].
A loss of cartilage function and quality is also seen with increasing age. There is a spectrum of diseases ranging from focal cartilage defects with healthy surrounding cartilage to focal lesions in degenerative cartilage, to multiple and diffuse lesions in osteoarthritic cartilage. Age-related factors such as senescent cells (SASP factors) may trigger tissue repair and remodelling by controlling cell reprogramming and dedifferentiation during wound healing. Cartilage microparticle transplantation, microfracture, intra-articular injection of drugs, joint washing and cleaning, total knee replacement, etc. are the traditional methods used to treat articular cartilage injury [6]. Regenerative techniques such as autologous chondrocyte transplantation (ACI) and matrix-induced autologous chondrocyte transplantation (MACI) have been applied in cartilage repair [7,8], but both are limited. Autologous grafts rely on cartilage or osteochondral plugs in non-weight-bearing areas of cartilage to fill cartilage defects, resulting in increased donor site lesions, in addition to potential donor site morbidity. On the other hand, allografts are complicated by immune rejection, potential disease metastasis and limited fusion with host tissue. Microfracture is mainly used for scar repair and involves disruption of subchondral bone to promote recruitment of multipotent bone marrow-derived stromal cells to the cartilage defect. However, this method usually leads to the formation of fibrocartilage, which has poor biological and mechanical properties, and may be painful for patients [9]. Intra-articular injection of drugs and joint washing and cleaning are suitable only for patients with early or mild disease, but their long-term effect is not ideal; a time-course analysis suggested that the efficacy of corticosteroid injections is reduced over time, and a meta-analysis of randomized clinical trials did not find a significant effect of intra-articular injections of hyaluronic acid (HA) compared with intra-articular injections of a placebo in the treatment of OA [10,11].
Growth factors are currently being considered in cartilage tissue engineering research. They have a wide range of functions and provide a new treatment for cartilage damage repair. Growth factors can regulate the synthesis and metabolism of the chondrocyte matrix, promote chondrogenic differentiation of stem cells, promote chondrocyte proliferation, maintain the chondrocyte phenotype and subsequently promote cartilage tissue regeneration, and repair articular cartilage damage. In addition to traditional growth factors, numerous new compounds that regulate cell growth, called newly discovered growth factors, are being reported, and some synthetic compounds and nonpolar particles also have differentiation-inducing effects on stem cells. Currently, among the latest developments in growth factor research, few studies have addressed the classification of growth factors suitable for stem cells from different sources. Thus, a greater understanding of these growth factors has great clinical significance for articular cartilage repair. In this study, we reviewed the roles of and researched progress on growth factors in cartilage injury and repair, starting from traditional research on growth factors and continuing to newly discovered growth factors and other synthetic compounds that can promote chondrogenic differentiation of stem cells.
Methodology
In this paper, the PubMed and SCI databases are searched with “artistic cartage”, “growth factor”, “stem cells” and “repair” as subject words. The “and” algorithm is used between the subject words, and a total of 1072 references are retrieved. Inclusion criteria: Basic research and clinical application of growth factors or cytokines in articular cartilage repair; 2. published from August 2000 to August 2019; 3. Select articles published in authoritative English journals in the same field, and select the core collection of papers by web of science. Exclusion criteria: 1. content irrelevant or not closely related research; 2. Non-English literature, repeatability study; 3. arguments, evidence unreliable literature; 4. Some documents that cannot be extracted from the data. Finally, 81 related articles with high quality were included, all of which were in English.
Results
A growth factor is essentially a kind of polypeptide substance. Growth factors can regulate cell function by binding with specific cell membrane receptors. They are also cytokines that can regulate cell growth. Growth factors can regulate the synthesis and metabolism of the chondrocyte matrix, promote chondrogenic differentiation of stem cells, promote chondrocyte proliferation, maintain the chondrocyte phenotype, promote cartilage tissue regeneration and repair articular cartilage damage. In the past few decades, traditional growth factors have been widely used in joint cartilage repair. In recent years, with the rapid development of articular cartilage repair, numerous new compounds that can regulate cell growth, called newly discovered growth factors, have been reported. In addition, some synthetic compounds and inorganic particles can induce chondrogenic differentiation of stem cells. Although many studies have addressed articular cartilage repair to date, the latest developments in growth factor research have not been evaluated. It is of great significance to fully understand the status of growth factor research for the treatment of articular cartilage repair defects (ACDs).
Bone morphogenetic proteins (BMPs)
BMPs belong to the transforming growth factor β (TGF-β) superfamily, which includes several other growth factors, such as activins, inhibins and TGFβs; BMPs are effective mediators of cell proliferation and MSC differentiation and have been suggested to be important molecules in cartilage repair [12]. BMPs can induce the formation of ectopic bone and cartilage, just as they mimic the formation of endochondral bone in embryos, and are important for maintaining bone integrity and fracture healing by inducing the differentiation of MSCs toward the osteoblast lineage. Currently, the BMP family contains approximately 20 members (BMP1-18, BMP⁃3b and BMP⁃8b) (Table 1) [13]. According to the similarity of the protein structure and gene homology, BMP family members can be subdivided into different subgroups, including the BMP- 2/4 subgroup, osteogenic protein-1 (OP-1) subgroup, more than 50% of the protein sequences in the BMP-9/10 subgroup, BMP- 12/13/14 subgroup, BMP-9/10 subgroup and BMP-12/13/14 subgroup. Bmp-11 and BMP-15 are distant members of the BMP family, similar to the growth and differentiation factors (GDF)- 8 and -9. Interestingly, the morphogenetic factors named GDFs are similar to some BMP family members; thus, they can also be included in the BMP family (Table 2). BMP-1, -2, -4, -5, -6, -7, -8, and -14 can induce the formation of bone and cartilage; BMP⁃2 and -7 can induce osteogenic/chondrogenic differentiation and bone and/or cartilage formation. BMP⁃2 can promote cartilage formation by chondrocytes and mesenchymal stem cells (MSCs) and is used to treat cartilage and cartilage interface injury [14,15]. BMP⁃3, 4, and 8 play key roles in the process of bone formation. BMP-3 can induce the proliferation of MSCs and promote the formation of cartilage [16], and BMP-3b can regulate cell growth and differentiation in embryonic and adult tissues [17]. BMP-4, 6, 7,-9, 12 and 13 has been shown to induce MSC differentiation. BMP-7 and -8 may be the inducers of epithelial osteogenesis, playing roles in calcium regulation and bone homeostasis and inducing cartilage and bone formation [14]. BMP-7 is used in cell carrier hydrogels to promote cartilage and extracellular matrix (ECM) formation [18]. When articular cartilage or bone marrow mesenchymal stem cells (BMSCs) were treated with BMP-7, the synthesis of type II collagen and mucopolysaccharide was promoted [19]. Iwakura et al. [20] showed that BMP-7 or TGF-β1 and BMP- 7 inhibited the differentiation of chondrocytes into mast cells in the early stage and effectively promoted synovial chondrogenesis. BMPs have a strong regulatory effect on the formation and repair of bone and cartilage, the proliferation of cells and the stability of the internal environment of adult bone. These molecules hold promise for articular cartilage repair and play a role in accelerating and increasing bone integration. Regeneration of the interface between full-thickness and osteochondral cartilage is a complex process involving repair of the biological functions of banded cartilage and subchondral bone. The mechanisms by which BMPs affect chondroprogenitor and osteoprogenitor cells to differentiate into cartilage or bone under different conditions need further study (Figure 1).
| BMP type | Nomenclature | Main physiological roles |
|---|---|---|
| Bone morphogenetic proteins | ||
| BMP-2 | BMP-2a | Cartilage and bone morphogenesis |
| BMP-3 | Osteogenin | Negative regulator of bone morphogenesis |
| BMP-3b | GDF-10 | Negative regulator of bone morphogenesis |
| BMP-4 | BMP-2b | Cartilage and bone morphogenesis |
| BMP-5 | - | Limb development/bone morphogenesis |
| BMP-6 | Vrg1, Dvr6 | Hypertrophy of cartilage/bone morphogenesis |
| BMP-7 | OP-1 | Cartilage and bone morphogenesis |
| BMP-8 | OP-2 | Bone morphogenesis/spermatogenesis |
| BMP-9 | GDF-2 | Bone morphogenesis/development of cholinergic neurons |
| BMP-11 | Axial skeleton patterning/eye development/pancreas development/kidney formation | |
| GDF-11 | ||
| Cartilage-derived morphogenetic proteins | ||
| BMP-12 | CDMP-3, GDF-7 | Ligament and tendon development |
| BMP-13 | CDMP-2, GDF-6 | Cartilage development and hypertrophy |
| BMP-14 | CDMP-1, GDF-5 | Chondrogenesis/angiogenesis |
Note: Growth and differentiation factor, GDF; osteogenic protein, OP.
Table 1: BMP members in humans and their main physiological roles (BMPs known to induce complete bone morphogenesis).
| Subgroup | Members of the TGFβ superfamily |
|---|---|
| BMP-2/4 | BMP-2, -4 |
| BMP-3/3b | BMP-3, BMP-3b (GDF-10) |
| OP-1 (BMP-7) | BMP-5, -6, -7, -8, -8b |
| BMP-9/10 | BMP-9 (GDF-2), BMP-10 |
| BMP-11/GDF-8 | BMP-11 (GDF-11), GDF-8 |
| BMP-15/GDF-9 | BMP-15 (GDF-9b), GDF-9 |
| CDMP1/2/3 | BMP-12, -13, -14 (GDF-5, -6, -7) |
Note: Cartilage-derived morphogenetic protein-CDMP.
Table 2: Suggested subgroups within the BMP family (grouped by similarity to OP-1).

Figure 1: Prisma flow diagram.
Cartilage-derived morphogenetic proteins (CDMPs)
The CDMP family consists of three members: CDMP⁃1, CDMP⁃2 and CDMP⁃3 [21]. They are highly similar in structure and biological characteristics; participate in the process of cartilage tissue formation, growth and damage repair; and play an important role in the differentiation and regulation of chondrocytes. CDMP-1 and CDMP-2 have 82% homology. Different species of CDMPs have high homology and low immunogenicity; thus, they generally do not cause immune rejection. Their low immunogenicity endows them with broad prospects for cross-species application [22]. In vitro experiments showed that CDMPs can promote the synthesis of proteoglycans from chondrogenic cell lines and fetal limb chondroblasts, indicating that CDMPs can stimulate the formation and differentiation of cartilage [23]. Cho et al. [24] showed that CDMPs can promote the production of glycosaminoglycan (GAG) by adult bovine and human articular chondrocytes, although the effect of CDMP-1 and CDMP-2 did not significantly differ. CDMP-1 is distributed mainly in the cartilage rudiment formed by MSC aggregation and the developing cartilage nucleus of long bone in human embryos, which are the sites of future joint formation; CDMP-1 is also expressed in adult articular cartilage. This specific distribution suggests a close relationship with the formation of articular cartilage in the extremities, while the distribution of articular cartilage in adults may be related to the maintenance of the chondrocyte phenotype. CDMP-1 can induce cartilage formation [25]. Tkahara et al. [26] found that some mesenchymal cells in the developing phalanx of short-legged rats lacking CDMP-1 appeared apoptotic and that the aggregation and differentiation of the cells in the finger (toe) were delayed when CDMP-1 was absent; thus, CDMP-1 was considered to be indispensable in the development, maintenance and growth of the finger (toe).
CDMP-1 can regulate the growth and differentiation of chondrocytes, maintain cellular phenotypes and promote apoptosis, and play specific roles in the differentiation of chondrocytes: (1) inducing the differentiation of stromal stem cells into chondrocytes; (2) promoting the aggregation and concentration of MSCs and inducing the differentiation of MSCs into chondrocytes; (3) determining the correct segmental pattern of the finger (toe) bone; and (4) maintaining the stable phenotype of mature articular chondrocytes; CDMP-1 can regulate the differentiation of MSCs into chondrocytes, which is closely related to chondrogenesis and development [27]. Spiro et al. [28] experimentally proved that CDMP-1 activity can be induced. For example, implantation of a matrix containing CDMP-1 into the subcutaneous tissue or muscle of mice can induce ectopic formation of cartilage and bone tissue. CDMP-1 can also promote the formation of cartilage nodules by cultured parietal cells of peptide-treated mice. Katayama et al. [29] transfected BMSCs with the CDMP-1 gene to repair ACDs in rabbits. Eight weeks after implantation of CDMP-1-transfected homologous BMSCs into ACDs in rabbits, the surface morphology of the repaired cartilage was similar to that of differentiated mature hyaline cartilage, and the subchondral tissue was completely repaired by the new thick layer of bone tissue near the host subchondral bone. The results of Bobacz et al. [30] showed that CDMP-1 can induce chondrogenic differentiation of fibroblasts. CDMP is also the inducer of osteogenesis in cartilage. CDMP-1 and CDMP-2 can promote the osteogenesis of BMSCs in vivo. Compared with growth or osteogenic medium alone, these proteins can dose-dependently increase the alkaline phosphatase activity.
TGF⁃β
TGF-β is a multifunctional polypeptide growth factor that exists widely in platelets and bone. TGF-β has many biological effects, such as regulating cell growth, differentiation, and apoptosis and extracellular matrix synthesis, and is the most potent cell growth factor identified to date. In addition, TGF⁃β can reduce chondrocyte hypertrophy and osteogenic differentiation, help promote the single process of chondrocyte differentiation during articular cartilage damage repair, and improve cartilage repair in vivo [31]. Five types of TGF-β have been discovered in humans, namely, TGF-β1, TGF-β2, TGF-β3, TGF-β4 and TGF-β5. They can effectively induce stem cells to differentiate into chondrocytes, and among the 5 types of TGF-β, TGF-β1 plays a more important role in cartilage damage repair than the other growth factors [32]. TGF-β 1 promotes cartilage damage repair in two ways: 1. it plays an inductive role, promoting stem cells toward chondrogenic differentiation; and 2. it promotes the synthesis of type II collagen and proteoglycan by chondrocytes, thus maintaining the chondrocyte phenotype. During mouse development, TGF-β1 mRNA is located in cartilage and the synovial intima, indicating that TGF-β1 promotes the growth and differentiation of cartilage tissue [33]. TGF-β1 can significantly increase cartilage-related gene expression and promote cell adhesion molecule expression and cell aggregation, while TGF β3 can promote cell proliferation and regulate chondrogenic differentiation of stem cells. TGF β1 and TGF β3 affect different stages of chondrogenic differentiation of stem cells. Therefore, the combined use of TGF-β1 and TGF-β3 may be better than either growth factor alone for inducing stem cells to differentiate into chondrocytes [34,35]. Li et al. [36] found that TGF-β not only induced chondrogenic differentiation but also inhibited the absorption of new tissue-engineered cartilage. In addition, TGF-β can regulate the expression and action of other cytokines in cartilage; for example, it can stimulate the synthesis of prostaglandins and type II collagen and can downregulate chondrodegradation enzymes. Additionally, TGF-β is an important synthetic metabolic factor in OA. It can protect cartilage from injury by counteracting the inhibitory effect of interleukin-1 (IL-1 ) on prostaglandins[37].
Fibroblast growth factors (FGFs)
FGFs can promote the division and proliferation of fibroblasts, induce the morphogenesis and differentiation of cells, and play an important role in the repair of bone and cartilage injury [38]. In different adult tissues, FGFs play an important role in the regulation of growth and tissue regeneration. Currently, FGF-2 and FGF-18 have been identified as having an important role in maintaining the homeostasis of articular cartilage, regulating chondrocyte differentiation and promoting the development of OA. FGF2 is derived from heparan sulfate proteoglycans in the extracellular matrix of articular cartilage, and its specific role in cartilage metabolism remains controversial [39]. According to related experimental studies, activation of the PI3K/AKT and ERK1/2 pathways by FGF2 can promote the proliferation of BMSCs, upregulate SOX9 gene expression, initiate the differentiation of BMSCs into cartilage chondrocytes, and promote the synthesis of extracellular matrix components such as type II collagen [40,41]. In a rabbit model of articular cartilage injury, the addition of exogenous FGF-2 significantly promoted the repair of injured articular cartilage. Argün et al. [42] successfully repaired full-thickness defects in rabbit articular cartilage by periosteal transplantation and injection of recombinant FGF-2. Shi et al. [43] stimulated the proliferation of adult bovine articular chondrocytes via FGF-2, which upregulated SOX9 and increased extracellular matrix production. FGF-1148 is a synthetic metabolic factor that can promote the proliferation of chondrocytes and participate in the formation of cartilage and repair of damaged cartilage. Zhang et al. [44] found that after coculture of chondrocytes from OA patients with MSCs and intervention with FGF-18, the cocultured cells could produce more type II collagen than OA chondrocytes cultured alone. FGF-18 was believed to promote the reconstruction and repair of damaged articular cartilage. In a recent study, Mori et al. [45]identified several Wnt proteins, insulin-like growth factors (IGFs), and TGF-β signaling-related molecules as predominant genes in articular cartilage. Previous genetic analysis studies showed that the expression of these factors including Dkk3, Wif1, Sfrp5, Igfbp4/5, Nov, Gdf10, Grem1 and Frzb in Articular chondrocytes was higher than that in growth plate chondrocytes.
Because articular cartilage cells are exposed to these factors under physiological conditions in the human body, they may respond better to these factors than to other factors. Therefore, researchers speculate that these signaling molecules may be more suitable than other molecules for joint cartilage tissue engineering. Moreover, related genetic analysis studies have shown that FGF-18 is an effective molecule that protects or regenerates adult articular cartilage. FGF-18 plays a very important role in bone growth and development because mice lacking FGF-18 show a variety of deformities, such as delayed closure of skull sutures, enlarged growth plates, and osteogenic differentiation disorders. Endogenous FGF-18 is abundantly expressed in the mature chondrocytes of articular cartilage, suggesting cell autonomy or autocrine/paracrine effects. Mori et al. [46] found that recombinant FGF-18 may be affected by the tissue inhibitor metalloproteinase-1 (TIMP-1) that protects articular cartilage through gene expression profile analysis. Howard et al. [47] injected recombinant human FGF-18 (rhFGF-18) into the articular cavity of sheep with medial femoral condyle defects and found that the articular cartilage was satisfactorily repaired. Through in vitro studies, Barr et al. [48] found that in addition to alterations in extracellular matrix macromolecules, an increase in FGF-18 accompanied the appearance of repair cells on the cartilage surface. Repair cells, characterized as chondrocytes, are observed on the surface of damaged articular cartilage and may participate in the repair response; their appearance requires both chondrocyte proliferation and migration of the cells to the damaged surfaces. The appearance of increased numbers of repair cells on damaged cartilage surfaces in the presence of rhFGF-18 following squamous intraepithelial lesion (SIL) injury suggests that FGF-18 influences cartilage repair via endogenous cell-mediated mechanisms in addition to affecting the extracellular matrix. In addition, recombinant FGF-18 significantly increased the synthesis of proteoglycan; thus, these researchers believed that FGF-18 can restore the number of chondrocytes and prevent chondrocyte apoptosis, which plays an important role in promoting the repair of mechanically damaged articular cartilage. FGFs increase the level of SOX-9 expression and enhance the activity of SOX-9-dependent chondrocyte-specific enhancer elements in the type II collagen gene [49]. Multiple signaling factors interact in a well-balanced manner to promote chondrogenesis; FGF has been shown to interact during chondrogenesis. Recently, the TGF-β, FGF, and Wnt protein families were demonstrated to control different stages of differentiation during chondrogenesis by crosstalk signaling [50].
IGFs
IGFs are single-chain polypeptides; their structure is homologous to that of insulin, and they are regulated by growth hormone. IGF was one of the first growth factors found to have a chondrogenic effect. IGF plays an important role in maintaining the homeostasis of chondrocytes and maintaining the balance of proteoglycan synthesis and decomposition in vitro. IGF in blood and synovial fluid can maintain the replacement of old and new chondrocytes. Loss of IGF abolishes the replacement of old and new chondrocytes. The IGF family has two members: IGF⁃1, which mainly regulates growth and development in adults, and IGF⁃2, which plays an important role in fetal growth and development. In normal articular cartilage, IGFs play a role in maintaining chondrocyte metabolic homeostasis and the balance of proteoglycan synthesis and decomposition in vitro. IGF-1 can promote chondrocyte proliferation, induce chondrogenic differentiation of MSCs, maintain cartilage phenotype stability, and promote articular cartilage repair. A study by Orth et al. [51]showed that transplantation of an alginate hydrogel coated with articular chondrocytes into rabbits promoted high expression of IGF-1. At 14 weeks, IGF1 improved articular cartilage regeneration, promoted the formation of subchondral bone, and enhanced the repair of full-thickness cartilage damage and osteochondral surface damage. Madry et al. [52] showed that overexpression of IGF-1 can significantly promote the repair of damaged cartilage and reduce the degeneration of cartilage around damaged areas in OA. In addition, Lu et al. [53] experimentally determined that increasing the content of IGF-1 in bilayered oligo(poly(ethylene glycol) fumarate) (OPF) hydrogel composites can promote cartilage formation. Loffredo et al. [54] implanted a collagen membrane loaded with IGF-1 into the cartilage defects in the right knee joints of rats. Low-dose IGF-1 promoted the repair and reconstruction of subchondral bone, and high-dose IGF-1 facilitated cell survival and promoted the new cartilage formation and cartilage integrity. IGFs can promote the proliferation of chondrocytes, induce chondrogenic differentiation of stem cells, stimulate cells to secrete extracellular matrix components, inhibit extracellular matrix-degrading enzymes, and then promote articular cartilage repair [55].
Kartogenin (KGN)
Johnson et al. [56] was the first to identify KGN, a small molecule that can enhance the differentiation of human mesenchymal stem cells (hMSCs) into chondrocytes. In addition, KGN can significantly promote the mRNA expression of TIMP-Ⅲ and collagen type II, and it can concentrate proteoglycan in human BMSCs, indicating that it has a strong ability to promote cartilage differentiation and can effectively promote the differentiation of MSCs into soft bone cells. Their research showed that KGN not only maintains the chondrocyte phenotype but also protects the cartilage matrix from degradation. Under simulated pathophysiological conditions in vitro, KGN can significantly reduce the level of chondrocyte-produced nitric oxide and the level of GAGs secreted by cartilage explants, suggesting that KGN has cartilage-protective effects on chondrocytes under pathological conditions. Yohei et al. [57] found that KGN inhibited the collapse of the extracellular matrix and aggrecan mediated by IL-1β and aggrecanase, stabilized the homeostasis of hyaluronic acid and CD44, and protected cartilage from injury. Xu et al. [58] established a model of rabbit articular cartilage injury and injected KGN into the cartilage defect, proving that KGN can enhance the regeneration of articular cartilage. Studies by Decker et al. [59] showed that KGN not only promotes the formation of cartilage but also enhances the development of limbs and joints in mice. Kwon et al. [60] showed that KGN can maintain the cartilage phenotype and prevent destruction of the extracellular matrix by matrix metalloproteinases (MMPs), indicating that KGN not only functions to repair articular cartilage but also has a protective effect on the original articular cartilage. Liu et al. [61] showed that KGN has a potent ability to promote chondrogenic differentiation of BMSCs and synovium⁃derived mesenchymal stem cells (SMSCs) and the repair of cartilage injury by stem cells. In addition, Liu et al. [62]found that KGN, TGF-β1 and BMP 7 can synergistically promote the secretion of lubricin by chondrocytes.
Platelet⁃rich fibrin (PRF) and platelet⁃rich plasma (PRP)
PRF and PRP are platelet concentrates prepared from whole blood by different centrifugation methods. They are rich in growth factors, including platelet-derived growth factor (PDGF), IGF, bFGF, TGF-β and vascular endothelial growth factor (VEGF), which can promote fracture healing, chondrocyte proliferation and cartilage extracellular matrix synthesis [63]. PRP can promote the synthesis of cartilage tissue at the defect site by promoting the proliferation of chondrocytes and the synthesis of cartilage-related matrices. Miyakoshi et al. [64] implanted the IGF-1-hyaluronic acid complex in PRP in the defect in a rabbit model and found that the complex promoted cartilage anabolism and repaired cartilage defects. Akeda et al. [65] cultured sheep chondrocytes in 10% PRP for 72 hours and found that the number of chondrocytes was significantly higher than that in cultures with normal culture medium. Lee et al. [66] added PRP combined with a chondrocyte/hydrogel scaffold to the culture medium of rabbit chondrocytes and found that PRP can promote chondrocyte proliferation, repair bone defects and promote cartilage formation. In a prospective study by Siclari et al. [67] 52 patients with knee joint cartilage injury were treated with a combination of subchondral drilling and PRP, and follow-up showed that the prognosis of these patients was good. Cytokines are released from PRP preparations at higher levels in early stages after use than in later stages; in addition, their release is temporary, uneven, and occurs mainly near the time point of exogenous additive supplementation. However, the pattern of cytokine release from PRF preparations is different from that from PRP preparations. Because PRF has no exogenous additives, the release of cytokines from its products is relatively long-lasting and more consistent with clinical needs.
PDGFPDGF is a large glycoprotein polypeptide growth factor that is synthesized, stored, and released mainly in the α particles of platelets. PDGF is a dimeric glycoprotein with a structure of two identical subunits (PDGF-AA, PDGF-BB) or two different subunits (PDGF-AB). The PDGF family contains five members: PDGF-AA, PDGF-BB, PDGF-AB, PDGF-CC and PDGF-DD. PDGF-BB is considered the most effective subtype in wound healing [68]. PDGF promotes cell proliferation and division, and the formation of extracellular matrix through contact with PDGF receptors on target cell membranes. Other cells, such as smooth muscle cells, endothelial cells, and activated macrophages, can also produce PDGF subtypes and release PDGF. In addition, PDGF plays an important role in the proliferation of osteoblasts, MSCs, tendon cells, fibroblasts, and vascular smooth muscle cells. Judith et al. [69] showed that PDGF can induce not only chemotactic macrophages but also fibroblasts and smooth muscle cells to migrate to the site of injury. Moreover, as a mitogen, PDGF promotes fibroblast proliferation and synthesizes extracellular matrix components such as GAGs, proteoglycans and collagen. Pujol et al. [70] confirmed that PDGF injection can increase the activity of meniscus cells and promote cartilage repair. Yokoyama et al. [71] collected rabbit platelet-activated serum (PAS), which is rich in PDGF BB. PAS was injected into the knee joints of Japanese White rabbits with anterior cruciate ligament transection, and the cartilage structure in all experimental groups was significantly improved compared with that in the control group.
VEGFVEGF is the strongest cytokine that can promote the proliferation of vascular endothelial cells. VEGF can increase the number of local blood vessels and improve local blood microcirculation. In addition, VEGF is an essential factor in the process of cartilage bone internalization. Through its role in promoting angiogenesis, it is involved in cartilage growth and development, bone repair after fracture, bone necrosis and bone metastasis of malignant tumors [72,73]. VEGF is related to the survival, differentiation, maturation, migration and absorption of bone (cartilage) osteoclasts [74]. Studies have shown that VEGFR-1 plays a very important role in stimulating the process of osteoclast production, and VEGFR-2 seems to have a greater effect on the stimulation of mature osteoclast function [75,76]. VEGF is also believed to indirectly stimulate the occurrence of osteoclasts by stimulating the production of RANKL in articular chondrocytes, osteoblasts and synovial fibroblasts. VEGF stimulates the migration, activity and differentiation of osteoblasts [74,76,77]. Both VEGFR-1 and VEGFR-2 participate in the osteogenic differentiation of MSCs through VEGF-mediated Runx2 expression [74]. VEGF activation of VEGFR-1 can induce osteoblast migration, and VEGFR-1 is the main receptor that mediates the biological effects of VEGF signaling on mature osteoblasts [77]. Clinical studies have evaluated anabolic growth factors, such as BMP-7/OP-1 or PRP, which contain a combination of growth factors and cytokines for the treatment of OA [78]. In animal models, growth factors, such as OP-1/BMP7 and BMP-4, and growth factor combinations, such as PRP, act synergistically with anti-VEGF or anti-angiogenesis therapies to promote cartilage repair and reduce the progression of OA [79,80]. Local application of anti-VEGF or antiangiogenic treatments at joint cartilage defects can improve therapeutic efficacy, reduce the number of injections, increase the safety, and enhance the clinical application value of the treatment. Methods to extend the release and duration of anti-VEGF and antiangiogenic therapies have been used in animal models of OA, as well as to enhance cartilage repair [80,81]. Considering that PRP contains VEGF, studies have shown that combining PRP with anti-VEGF antibodies can limit the potential negative effects of PRP in the treatment of OA or cartilage defects while still allowing the potential benefits of other growth factors and cytokines [82].
Epidermal growth factor (EGF)
EGF is a polypeptide consisting of 53 amino acid residues, with a molecular weight of 6045 kD. The structure of EGF contains 3 intramolecular disulfide bonds, which are the basis of its biological activity and have a proliferative effect on a variety of cells. EGF can cause epithelial cells of various tissues to undergo mitosis and can stimulate the proliferation and differentiation of mesenchymal cells. Studies have shown that EGF receptors are expressed on osteoblasts, osteoclasts and endothelial cells [83]. Wang et al. [84] cultured rabbit articular chondrocytes with EGF and VEGF in vitro and found that EGF can promote cell survival and proliferation and that the combination of EGF and IGF had the optimal synergistic effect. However, Nonaka K et al. [85] found that EGF and BMP-4 play antagonistic roles in cartilage differentiation; BMP-4 plays a promotive role, but EGF plays a suppressive role, and both cytokines act on cartilage through Smad l. Research by Huh et al.[86]showed that the EGF exerts its inhibitory effect by stimulating the expression of COX-2 and PGE-2 through the ERK and p38 kinase signaling pathways to inhibit the synthesis of type II collagen and proteoglycans. Janssen et al. [87] found that EGF, as well as the osteochondrogenic regulators SOX9 and RUNX2, was expressed in articular cartilage. The direct stimulation of chondrogenic progenitor cells (CPCs) in vitro with TGF-β3 and EGF increased the level of the chondrogenic master regulator SOX9 but also increased the levels of osteogenic proteins and hypertrophy markers.
Nerve growth factor (NGF)
Studies have shown that NGF levels in the joints of patients with osteoarthritis are increased and are involved in OA-related pain [88]. Research by Pecchi et al. found that the articular chondrocytes of unstimulated mice and humans express low levels of NGF [89]. NGF stimulates the growth of sensory nerves, and the expression of NGF increases in the inflamed synovium. NGF can also stimulate angiogenesis, indicating that there is a close connection between blood vessels and nerve growth [90]. Biglycan (BGN) interacts with components of TGF-β3 and EGF pathways and binds to mediators and receptors to increase its activity. Both TGF-β3 and EGF signaling pathways affect chondrocyte differentiation [91]. Janssen et al. [87] found that EGF, TGF and BGN are expressed in articular cartilage and may affect each other's expression and the expression of osteogenic regulators RUNX2 and SOX9. Direct stimulation of CPCs cultured in vitro with EGF and TGF-β3 can increase the level of SOX9, the master regulator of chondrogenesis, and also increase hypertrophy markers and osteogenic capacity.
Nerve growth is accompanied by neovascularization in various tissues, including subcutaneous cavernoma and healing fractures in animal models. NGF sensitizes nerves; therefore, increased expression of NGF in OA may increase sensory nerve activity in subchondral bone [92].
WNT signaling
The WNTs are a family of 19 morphogens that play fundamental roles in developmental processes ranging from embryonic morphogenesis to homeostasis of adult tissues; more than 20 WNT family members are found in vertebrates [93]. The WNT pathway is divided into at least two branches: β-catenin-independent “non-canonical” signaling pathways and canonical WNT/β-catenin signaling pathways [94]. To regulate the expression of various genes, WNT family members are activated by at least four different b-catenin independent pathways; that is, the protein kinase C, PLC, and calcium/calmodulin-dependent protein kinase II and JNK pathways [95]. Different WNT family members play important roles in cartilage formation and bone development, among which Wnt-1, Wnt-4, Wnt-7a, and Wnt-8 can prevent cartilage differentiation, despite exhibiting different effects during hypertrophy; however, Wnt5a, Wnt-5b, and Wnt-11 regulate the proliferation and hypertrophy of growth plate chondrocytes during development [96]. Most cellular effects of WNT signal transduction influence the regulation of chondrogenesis and the dedifferentiation of primary articular chondrocytes through the modulation of β-catenin. For example, WNT-3A and WNT-7A stimulate β-catenin-mediated transcription [97,98]. Accumulation of β-catenin and subsequent activation of the β-catenin-TCF/LEF complex induces the dedifferentiation of articular chondrocytes by inhibiting the expression of COL-2 and activating the expression of COL-1; in addition, Wnt-3A induces the c-Jun expression and c-Jun-mediated phosphorylation of JNK, leading to ap-1 activation, thereby inhibiting the expression of SOX-9 [98].Typical WNT signalling inhibits cartilage formation and induces ossification, although the effects of Wnt-3A on cartilage formation are controversial [99]. In adult tissues, cartilage formation and the osteogenic process require β-catenin [100]. In general, WNT plays an important role in cartilage. Developmental regulation of chondrocytes and an imbalance in WNT networks may lead to arthritis, especially OA. The WNT signaling pathway modulates key biological processes in development, growth, homeostasis, and disease, particularly in joints and bone. Excessive activation of WNT signaling pathways in articular cartilage has been clearly associated with the onset and severity of OA. Hence, targeting WNT signaling may be an excellent approach to develop specific drugs that may be useful in the treatment of OA [92, 94, 98-100].
Mechano-growth factor (MGF)MGF is an alternative splicing product of IGF-1. MGF contains only 24 peptides, which are formed from IGF-1 splice variants of exon 4 spliced to exon 5 (or exon 6), and is named MGF (or IGF-IEc) in humans and IGF-IEb in rodents. MGF and IGF are regulated and expressed by the same gene and are isomers of the IGF family, but their physiological functions differ greatly [101]. Tong et al. [102] showed that MGF can regulate the migration of MSCs through the PI3K/AKT and Erk1/2 signaling pathways and promote the proliferation and osteogenic differentiation of MSCs. Luo et al. [103] demonstrated that MGF is a catalyst for promoting TGF-β3-induced cartilage regeneration. In animal experiments, fibroin scaffolds carrying TGF-β3 and MGF induce BMSC migration during the repair of ACDs in rabbits and inhibit the formation of fibrosis on the cartilage surface, which can promote regeneration and repair of articular cartilage over an extended period. Deng et al. [104] injected MGF (28.5 mg/kg and 57 μg/kg) into areas of bone deformation in rabbits to promote osteogenesis and proved that high-dose MGF was more effective than low-dose MGF. In vitro, MGF significantly promotes the migration and TGF-β3-mediated chondrogenic differentiation of BMSCs and reduces the synthesis of type I collagen during cartilage differentiation. The surface of the new cartilage is smooth and flat, and the matrix is rich in proteoglycan. Although the new cartilage does not express type I collagen, it can express normal type II collagen, and the repair effect is very similar to that of normal cartilage. Song et al. [105] showed that MGF can stimulate anabolism in the OA process and that MGF can upregulate the cartilage extracellular matrix synthesis genes Col II, ACAN, SOX9 and HAS. With increasing MGF dose, the expression of Col II, ACAN and Sox9 increased linearly, indicating that MGF has a stable effect on extracellular matrix regeneration and cartilage formation.
Inorganic particles
Various types of hydrogels derived from different natural or synthetic polymers and their hybrids have been used to reconstruct defective osteochondral interfaces or articular cartilage tissue [106]. Hydrogel complexes containing inorganic particles are new functional materials used for the treatment of osteochondral surfaces and full-thickness cartilage damage. Inorganic particles can enhance the mechanical properties of hydrogels, and they have pronounced bone conductivity properties and osteogenic induction ability [107,108]. Khanarian et al. [109]added hydroxyapatite nanoparticles into agarose hydrogels, which increased the total amount of proteoglycan, collagen and calcification and promoted regeneration of the bone cartilage interface and calcified cartilage. Han et al. [107] added calcium silicate microparticles to alginate hydrogels to stimulate osteogenic differentiation of MSCs and significantly increase the deposition of extracellular bone-like minerals. Filardo et al. [110] treated 27 patients with osteochondritis dissecans with a three-dimensional permeable scaffold composed of type I collagen and hydroxyapatite before surgery; cartilage defects were exposed, and sclerotic subchondral bone was removed via open surgery. After 2 years of follow-up, magnetic resonance imaging (MRI) showed that cartilage defects were repaired, and most patients exhibited morphological changes in the subchondral bone and uneven tissue regeneration.
Dexamethasone
Dexamethasone is an adrenocortical hormone often used to treat OA caused by articular cartilage injury, and it has been proven to promote chondrogenesis and osteogenic differentiation of stem cells [111]. Adding dexamethasone to culture medium or covalently binding it to a hydrogel can induce MSCs to differentiate into osteoblasts, thereby improving osteogenesis [112]. Florine et al. [113] showed that in the presence of TGF-β, dexamethasone can enhance chondrogenic differentiation of MSCs and ESCs and the formation of chondro-related proteins. Dexamethasone is usually used to supplement TGF or BMP to further promote cell proliferation and maximize chondrogenesis or osteogenic induction, thus achieving optimal chondrogenesis or osteogenic effects [114].
MSCs are the most widely used stem cells in regenerative medicine. Because of their abundant sources, low immunogenicity, and lack of ethical issues [115], they can be encapsulated in various hydrogels; therefore, MSC carrier hydrogels have become the most promising biomaterials for cartilage tissue engineering and can be used for osteochondral interface regeneration and cartilage repair. The abilities of hydrogels to support cartilage formation and osteogenesis vary. Histological and immunohistochemical staining have confirmed that MSCs form cartilage and bone extracellular matrix after culture in alginate, chitosan and silk protein hydrogels for 8 weeks; however, the differentiation of MSCs is multidirectional. Specific cell growth factors must be added to correctly induce MSCs to differentiate into chondrocytes. In addition, stem cells from different sources and different types of hydrogels show different abilities to support cartilage and bone formation, and different growth factors are needed to induce stem cells to differentiate in to chondrocytes (Table 3).
| Types | Families of growth factors | Family members | Commonly used family members | Applicable stem cells | Animal models | Clinical experiments |
|---|---|---|---|---|---|---|
| Bone morphogenetic proteins(BMPs) | BMP-1-18, BMP-3b, BMP-8b | BMP-2 | BMSCs, ESCs, | Rabbit femoral condyle, Rat femoral condyle | - | |
| BMP-7 | ADSCs, SMSCs | |||||
|
|
|||||
|
|
|||||
| Cartilage-derived morphogenetic proteins(CDMPs) | CDMP-1, CDMP-2, CDMP-3 | CDMP⁃1 | BMSCs, ADSCs | Rabbit femoral condyle | - | |
| Transcription growth factor -β (TGF-β) | TGF⁃β1, TGF⁃β2, TGF⁃β3, TGF⁃β1β2, TGF⁃β4, TGF⁃β5 | TGF⁃β1, | BMSCs, PSCs, | Rabbit femoral condyle, Rat patella, Human osteochondral model | ||
| TGF⁃β3 | ESCs, ADSCs | - | ||||
| Traditional growth factors | Fibroblast growth factors (FGFs) | aFGF, bFGF | bFGF(FGF⁃2, FGF⁃18) | BMSCs | Rabbit femoral condyle, Rat OA model | - |
|
Insulin⁃like growth factors(IGFs) | IGF⁃1, IGF⁃2 | IGF⁃1 | BMSCs, ADSCs | Rabbit femoral condyle | - |
| Kartogenin (KGN) | KGN | KGN | BMSCs, SMSCs | Rat OA model; Rabbit femoral condyle | - | |
| Newly discovered cell growth factors | Platelet concentrate | PRP, PRF | PRP, PRF | BMSCs, ADSCs, | Rabbit femoral condyle | + |
|
SMSCs | |||||
|
Mechano⁃growth factor (MGF) | MGF | MGF | BMSCs | Rabbit femoral condyle | - |
| Inorganic particles | Phosphate compounds, silicate compounds, etc. | Phosphate compounds, silicate compounds, etc. | BMSCs, SMSCs | - | - | |
| Synthetic compounds | Dexamethasone | Dexamethasone | Dexamethasone | BMSCs, ESCs, | - | - |
|
ADSCs, SMSCs |
Note: ESCs, Embryonic Stem Cells; PSCs, Pluripotent Stem Cells; SMSCs, Synovium-derived Mesenchymal Stem Cells; ADSCs, Adipose-derived Stem Cells; BMSCs, Bone marrow Mesenchymal Stem Cells.
Table 3: Some common cell growths in cell repair.
Discussion
In articular cartilage, both acute injury and slow progression of OA cause pain, leading to progressive destruction of the whole joint, resulting in partial or complete loss of joint function. Articular cartilage injury repair is an extremely complex process. Clinical treatments currently used to repair cartilage defects are palliative and not curative. Although chondrocyte transplantation and early tissue engineering techniques have achieved some success, the regenerative ability of cartilage tissue is limited. The use of biological methods to regenerate cartilage is still a great challenge. Growth factors have a wide range of effects and provide a new treatment approach for cartilage damage repair, a popular focus of cartilage tissue engineering research. Chondrocytes inhabit a specific "cartilage microenvironment" with complex and diverse internal components, not simply a single growth factor; in the "cartilage microenvironment", multiple growth factors cooperate to enhance cartilage matrix synthesis and promote cartilage repair. In articular cartilage, multiple growth factors enhance the healing ability of cartilage injury and prevent the occurrence and development of degenerative arthritis. To induce chondrocyte differentiation, it is necessary to stimulate and promote chondrocyte differentiation with appropriate stimulatory factors. Research has proven that many growth factors and preparations, including BMPs, CDMPs, TGF-βs, IGF-1, FGFs, HGF, KGN, PDGF, MGF, PRP, and PRF, can induce or stimulate chondrocyte differentiation. Moreover, recent studies have shown that the effect of multi-growth factor combinations is significantly greater than that of a single factor (Mendes et al. 2018). Previous studies focused on traditional growth factors, and many studies have studied the role of single growth factors; however, few studies have focused on the combined application of different growth factors. The synergistic or antagonistic effects of different growth factors, the differences between their in vivo and in vitro effects, and the optimal concentration ratios need further study. To date, most research on growth factors has been basic research, and few clinical trials and clinical studies have been conducted. Thus, clinical research is the next key research direction. No experimental study on the use of dexamethasone and inorganic particles in vivo has been conducted to date, and further study is required on the repair effect and safety of dexamethasone and inorganic particles in vivo. The research on growth factors described in this paper may help to guide the combined application of multiple growth factors for articular cartilage repair. However, the crucial problem in the combined application of multiple growth factors is to determine the optimal combinations; thus, researchers should consider the synergistic effect of multiple growth factors on cartilage repair.
Conclusion
Despite the continuous progress in surgical technologies, the treatment of cartilage injury is still a considerable clinical challenge. Although the newly discovered growth factors have greatly promoted the development of articular cartilage repair approaches, and some of the results have been applied clinically, their specific mechanisms are unclear. To date, researchers have focused more on the effects of growth factors on chondrocyte proliferation and morphogenesis than on the signaling pathways that affect growth factors. Through in-depth research on the roles of various growth factors in cartilage damage, the potential of growth factors in cartilage repair will continue to be further explored. Approaches for measuring the levels of specific growth factors, the interactions between growth factors and the healing environment, and the regulatory mechanism of growth factors in the differentiation of repair cells need further clarification to improve the application of growth factors, ensure the use of optimal timing and dosages during the cartilage defect repair process, and provide a basis for identifying joint lesions. Through in-depth study of the influence of interactions between various growth factors on the cartilage repair process and the influence of external factors on the "cartilage microenvironment", approaches to use growth factors alone or in combination to treat cartilage tissue damage can be developed.
Declarations
Ethics approval and consent to participate
Not applicable
Consent for Publication
Not applicable
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing Interests
The authors declare that they have no competing interests.
Funding
Not applicable
Authors' Contributions
Yu Zhang designed this paper to analyze and interpret data about growth factors in articular cartilage repair. Zishu Chai performed the data collection, and Chengqiang Yu, Youcai Wu and Yufu Ou were responsible for the literature analysis. Yu Zhang was responsible for and was the main contributor to writing the manuscript. Jianxun Wei was responsible for proofreading, and Yu Zhang was responsible for the final version of the article. All authors read and approved the final manuscript.
Acknowledgments
Not applicable
REFERENCES
- Neogi T, Zhang Y. Epidemiology of osteoarthritis. Rheum Dis Clin. 2013;39(1):1-9.
- Bitton R. The economic burden of osteoarthritis. Am J Manag Care. 2009;15:230-235.
[Google Scholar] [PubMed]
- Tuan RS, Chen AF, Klatt BA. Cartilage regeneration. J Am Acad Orthop Surg. 2013;21(5):303.
- Hunziker EB, Lippuner K, Keel MJ, Shintani N. An educational review of cartilage repair: Precepts & practice-myths & misconceptions-progress & prospects. Osteoarthr Cartil. 2015;23(3):334-350.
[Crossref] [Google Scholar] [PubMed]
- Mithoefer K, McAdams T, Williams RJ, Kreuz PC, Mandelbaum BR. Clinical efficacy of the microfracture technique for articular cartilage repair in the knee: An evidence-based systematic analysis. Am J Sports Med. 2009;37(10):2053-2063.
[Crossref] [Google Scholar] [PubMed]
- Ye K, Di Bella C, Myers DE, Choong PF. The osteochondral dilemma: Review of current management and future trends. ANZ J Surg. 2014;84(4):211-217.
[Crossref] [Google Scholar] [PubMed]
- Dewan AK, Gibson MA, Elisseeff JH, Trice ME. Evolution of autologous chondrocyte repair and comparison to other cartilage repair techniques. BioMed Res Int. 2014.
[Crossref] [Google Scholar] [PubMed]
- Kon E, Filardo G, Di Martino A, Marcacci M. ACI and MACI. J Knee Surg. 2012;25(01):017-22.
- Mithoefer K, McAdams T, Williams RJ, Kreuz PC, Mandelbaum BR. Clinical efficacy of the microfracture technique for articular cartilage repair in the knee: An evidence-based systematic analysis. Am J Sports Med. 2009;37(10):2053-2063.
[Crossref] [Google Scholar] [PubMed]
- Jüni P, Hari R, Rutjes AW, Fischer R, Silletta MG, Reichenbach S, da Costa BR. Intraâ?articular corticosteroid for knee osteoarthritis. Cochrane Database Syst Rev. 2015(10).
- Jevsevar D, Donnelly P, Brown GA, Cummins DS. Viscosupplementation for osteoarthritis of the knee: A systematic review of the evidence. JBJS. 2015;97(24):2047-2060.
[Crossref] [Google Scholar] [PubMed]
- Carreira AC, Alves GG, Zambuzzi WF, Sogayar MC, Granjeiro JM. Bone Morphogenetic Proteins: Structure, biological function and therapeutic applications. Arch Biochem Biophys. 2014;561:64-73.
[Crossref] [Google Scholar] [PubMed]
- Liu M, Zeng X, Ma C, Yi H, Ali Z, Mou X, et al. Injectable hydrogels for cartilage and bone tissue engineering. Bone res. 2017;5(1):1-20.
[Crossref] [Google Scholar] [PubMed]
- López-Ruiz E, Jiménez G, Kwiatkowski W, Montañez E, Arrebola F, Carrillo E, et al. (2018) Impact of TGF-β family-related growth factors on chondrogenic differentiation of adipose-derived stem cells isolated from lipoaspirates and infrapatellar fat pads of osteoarthritic patients. Eur Cells Mater 35:209.
[Crossref] [Google Scholar] [PubMed]
- Taniyama T, Masaoka T, Yamada T, Wei X, Yasuda H, Yoshii T, et al. Repair of osteochondral defects in a rabbit model using a porous hydroxyapatite collagen composite impregnated with bone morphogenetic proteinâ?2. Artificial Organs. 2015;39(6):529-535.
[Crossref] [Google Scholar] [PubMed]
- Chen G, Deng C, Li YP. TGF-β and BMP signaling in osteoblast differentiation and bone formation. Int J Biol Sci. 2012;8(2):272.
[Crossref] [Google Scholar] [PubMed]
- Dimitriou R, Tsiridis E, Giannoudis PV. Current concepts of molecular aspects of bone healing. Injury. 2005;36(12):1392-1404.
[Crossref] [Google Scholar] [PubMed]
- Jose CC, Borrajo E, Vidal A. New scaffolds encapsulating TGFâ? β3/BMPâ?7 combinations driving strong chondrogenic differentiation. J Eur J Pharm Biopharm. 2017;114:69-78.
[Crossref] [Google Scholar] [PubMed]
- Krase A, Abedian R, Steck E, Hurschler C, Richter W. BMP activation and Wnt-signalling affect biochemistry and functional biomechanical properties of cartilage tissue engineering constructs. Osteoarthr Cartil. 2014;22(2):284-292.
[Crossref] [Google Scholar] [PubMed]
- Iwakura T, Sakata R, Reddi AH. Induction of chondrogenesis and expression of superficial zone protein in synovial explants with TGF-β1 and BMP-7. Tissue Eng. 2013;19(24):2638-2644.
[Crossref] [Google Scholar] [PubMed]
- Tran LP, Zalzal GH. Laryngotracheal stenosis repair using cartilage-derived morphogenic proteins: A pilot study. Ann Otol Rhinol Laryngol. 2006;115(6):477-481.
[Crossref] [Google Scholar] [PubMed]
- Miyamoto Y, Mabuchi A, Shi D, Kubo T, Takatori Y, Saito S, et al. A functional polymorphism in the 5â?² UTR of GDF5 is associated with susceptibility to osteoarthritis. Nat Genet. 2007;39(4):529-533.
[Crossref] [Google Scholar] [PubMed]
- Takahara M, Harada M, Guan D, Otsuji M, Naruse T, Takagi M, et al. Developmental failure of phalanges in the absence of growth/differentiation factor 5. Bone. 2004;35(5):1069-1076.
[Google Scholar] [PubMed]
- Cho TJ, Gerstenfeld LC, Einhorn TA. Differential temporal expression of members of the transforming growth factor β superfamily during murine fracture healing. J Bone Miner Res. 2002;17(3):513-520.
[Crossref] [Google Scholar] [PubMed]
- Guo CA, Liu XG, Huo JZ, Jiang C, Wen XJ, Chen ZR. Novel gene-modified-tissue engineering of cartilage using stable transforming growth factor-β1-transfected mesenchymal stem cells grown on chitosan scaffolds. J Biosci Bioeng. 2007;103(6):547-556.
[Crossref] [Google Scholar] [PubMed]
- Takahara M, Harada M, Guan D, Otsuji M, Naruse T, Takagi M, Ogino T. Developmental failure of phalanges in the absence of growth/differentiation factor 5. Bone. 2004;35(5):1069-1076.
[Crossref] [Google Scholar] [PubMed]
- Feng G, Wan Y, Balian G, Laurencin CT, Li X. Adenovirus-mediated expression of growth and differentiation factor-5 promotes chondrogenesis of adipose stem cells. Growth Factors. 2008;26(3):132-142.
[Crossref] [Google Scholar] [PubMed]
- Spiro RC, Liu LS, Heidaran MA, Thompson AY, Ng CK, Pohl J, et al. Inductive activity of recombinant human growth and differentiation factor-5.
[Crossref] [Google Scholar] [PubMed]
- Katayama R, Wakitani S, Tsumaki N, Morita Y, Matsushita I, Gejo R, et al. Repair of articular cartilage defects in rabbits using CDMP1 gene-transfected autologous mesenchymal cells derived from bone marrow. Rheumatology. 2004;43(8):980-985.
[Crossref] [Google Scholar] [PubMed]
- Bobacz K, Ullrich R, Amoyo L, Erlacher L, Smolen JS, Graninger WB. Stimulatory effects of distinct members of the bone morphogenetic protein family on ligament fibroblasts. Ann Rheum Dis. 2006;65(2):169-177.
[Crossref] [Google Scholar] [PubMed]
- Frisch J, Reyâ?Rico A, Venkatesan JK, Schmitt G, Madry H, Cucchiarini M. TGFâ?β gene transfer and overexpression via rAAV vectors stimulates chondrogenic events in human bone marrow aspirates. J Cell Mol Med. 2016;20(3):430-440.
[Crossref] [Google Scholar] [PubMed]
- Deng Y, Sun AX, Overholt KJ. Enhancing chondrogenesis and mechanical strength retention in physiologically relevant hydrogels with incorporation of hyaluronic acid and direct loading of TGF-β[J]. Acta Biomater. 2019;83:167-176.
[Crossref] [Google Scholar] [PubMed]
- Takahashi N, Rieneck K, Kraan PM. van der, Beuningen HM van, Vitters EL, et al. Elucidation of IL-1/TGF-Ã? interactions in mouse chondrocyte cell line by genomewide gene expression. Osteoarthr cartil. 2005;13:426-438.
[Crossref] [Google Scholar] [PubMed]
- Askari M, Bonakdar S, Anbouhi MH, Shahsavarani H, Kargozar S, Khalaj V, et al. Sustained release of TGF-β1 via genetically-modified cells induces the chondrogenic differentiation of mesenchymal stem cells encapsulated in alginate sulfate hydrogels. J Mater Sci: Mater Med. 2019;30(1):1.
- Duchi S, Doyle S, Eekel T, D Oâ??Connell C, Augustine C, Choong P, et al. Protocols for culturing and imaging a human ex vivo osteochondral model for cartilage biomanufacturing applications. Materials. 2019;12(4):640.
[Crossref] [Google Scholar] [PubMed]
- Li C, Bi W, Gong Y, Ding X, Guo X, Sun J, et al. Transforming growth factor-beta1 inhibits tissue engineering cartilage absorption via inducing the generation of regulatory T cells. Tissue Eng Regen Med. 2016;10(2):113-120.
[Crossref] [Google Scholar] [PubMed]
- Riera KM, Rothfusz NE, Wilusz RE, Weinberg JB, Guilak F, McNulty AL. Interleukin-1, tumor necrosis factor-alpha, and transforming growth factor-beta 1 and integrative meniscal repair: Influences on meniscal cell proliferation and migration. Arthritis Res Ther. 2011;13(6):1-20.
[Crossref] [Google Scholar] [Pubmed]
- Hu WQ, Zhao JN, Xu HD, et al. Research advances of growth factors in cartilage repair[J]. J Clin Pathol Res. 2015;35(3): 474â?478.
- Ornitz DM, Marie PJ. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015;29(14):1463-1486.
[Crossref] [Google Scholar] [PubMed]
- Correa D, Somoza RA, Lin P, Greenberg S, Rom E, Duesler L. Sequential exposure to fibroblast growth factors (FGF) 2, 9 and 18 enhances hMSC chondrogenic differentiation. Osteoarthr Cartil. 2015;23(3):443-453.
[Crossref] [Google Scholar] [PubMed]
- Yang W, Cao Y, Zhang Z, Du F, Shi Y, Li X. Targeted delivery of FGF2 to subchondral bone enhanced the repair of articular cartilage defect. Acta biomaterialia. 2018;69:170-182.
[Crossref] [Google Scholar] [PubMed]
- Argün M, �ner M, Güney A, Halıcı M, Temizyürek O, Canöz �. The healing of full-thickness articular cartilage defects in rabbits: successful results with fibroblast growth factor. Jt Dis Relat Surg. 2010;21(3):147-152.
[Google Scholar] [PubMed]
- Shi S, Wang C, Acton AJ, Eckert GJ, Trippel SB. Role of sox9 in growth factor regulation of articular chondrocytes. J Cell Biochem. 2015;116(7):1391-1400.
[Crossref] [Google Scholar] [PubMed]
- Zhang Zï¼?Wang Yï¼?Li M. Fibroblast growth factor 18 increases the trophic effects of bone marrow mesenchymal stem cells on chondrocytes isolated from late stage osteoarthritic patients. Stem Cell Int. 2014(2014):1-8.
[Crossref] [Google Scholar] [PubMed]
- Mori Y, Saito T, Chang SH, Kobayashi H, Ladel CH, Guehring H. Identification of fibroblast growth factor-18 as a molecule to protect adult articular cartilage by gene expression profiling. J Biol Chem. 2014;289(14):10192-10200.
[Crossref] [Google Scholar] [PubMed]
- Howard D, Wardale J, Guehring H, Henson F. Delivering rhFGFâ?18 via a bilayer collagen membrane to enhance microfracture treatment of chondral defects in a large animal model. J Orthop Res. 2015;33(8):1120-1127.
[Crossref] [Google Scholar] [PubMed]
- Barr L, Getgood A, Guehring H, Rushton N, Henson FM. The effect of recombinant human fibroblast growth factorâ?18 on articular cartilage following single impact load. J Orthop Res. 2014;32(7):923-927.
[Crossref] [Google Scholar] [PubMed]
- Murakami S, Kan M, McKeehan WL, De Crombrugghe B. Up-regulation of the chondrogenic Sox9 gene by fibroblast growth factors is mediated by the mitogen-activated protein kinase pathway. Proc Natl Acad Sci. 2000;97(3):1113-1118.
[Crossref] [Google Scholar] [PubMed]
- Cleary MA, van Osch GJ, Brama PA, Hellingman CA, Narcisi R. FGF, TGFβ and Wnt crosstalk: Embryonic to in vitro cartilage development from mesenchymal stem cells. J Tissue Eng Regen Med. 2015 ;9(4):332-342.
[Crossref] [Google Scholar] [PubMed]
- Orth P, Kaul G, Cucchiarini M, Zurakowski D, Menger MD, Kohn D, et al. Transplanted articular chondrocytes co-overexpressing IGF-I and FGF-2 stimulate cartilage repair in vivo. Knee Surg Sports Traumatol Arthrosc. 2011;19(12):2119-2130.
[Crossref] [Google Scholar] [PubMed]
- Madry H, Kaul G, Zurakowski D, Vunjak-Novakovic G, Cucchiarini M. Cartilage constructs engineered from chondrocytes overexpressing IGF-I improve the repair of osteochondral defects in a rabbit model. Eur Cells Mater. 2013;25:229.
[Crossref] [Google Scholar] [PubMed]
- Lu S, Lam J, Trachtenberg JE, Lee EJ, Seyednejad H, van den Beucken JJ, Tabata Y, et al. Dual growth factor delivery from bilayered, biodegradable hydrogel composites for spatially-guided osteochondral tissue repair. Biomaterials. 2014;35(31):8829-8839.
[Crossref] [Google Scholar] [PubMed]
- Loffredo FS, Pancoast JR, Cai L, Vannelli T, Dong JZ, Lee RT, et al. Targeted delivery to cartilage is critical for in vivo efficacy of insulinâ?like growth factor 1 in a rat model of osteoarthritis. Arthritis&rheumat. 2014;66(5):1247-1255.
[Crossref] [Google Scholar] [PubMed]
- Bessa PC, Casal M, Reis RL. Bone morphogenetic proteins in tissue engineering: the road from the laboratory to the clinic, part I (basic concepts). J Tissue Eng Regen Med. 2008;2(1):1-3.
[Crossref] [Google Scholar] [PubMed]
- Johnson K. Based Approach to Cartilage Repair-A Stem Cell. Sci. 2012;1215157(717):336.
- Ono Y, Ishizuka S, Knudson CB, Knudson W. Chondroprotective effect of kartogenin on CD44-mediated functions in articular cartilage and chondrocytes. Cartilage. 2014;5(3):172-180.
[Crossref] [Google Scholar] [PubMed]
- Xu X, Shi D, Shen Y, Xu Z, Dai J, Chen D, et al. Full-thickness cartilage defects are repaired via a microfracture technique and intraarticular injection of the small-molecule compound kartogenin. Arthritis Res Ther. 2015;17(1):1-2.
[Crossref] [Google Scholar] [PubMed]
- Decker RS, Koyama E, Enomoto-Iwamoto M, Maye P, Rowe D, Zhu S, et al. Mouse limb skeletal growth and synovial joint development are coordinately enhanced by Kartogenin. Dev Biol. 2014;395(2):255-267.
[Crossref] [Google Scholar] [PubMed]
- Kwon JY, Lee SH, Na HS, Jung K, Choi J, Cho KH, et al. Kartogenin inhibits pain behavior, chondrocyte inflammation, and attenuates osteoarthritis progression in mice through induction of IL-10. Sci Rep. 2018;8(1):1-1.
[Crossref] [Google Scholar] [PubMed]
- Liu C, Li T, Yang Z, Liu D, Li Y, Zhou Z, Zhang Q. Kartogenin enhanced chondrogenesis in cocultures of chondrocytes and bone mesenchymal stem cells. Tissue Engineering Part A. 2018;24(11-12):990-1000.
[Crossref] [Google Scholar] [PubMed]
- Liu C, Ma X, Li T. Kartogenin, transforming growth factor-β1 and bone morphogenetic protein-7 coordinately enhance lubricin accumulation in bone-derived mesenchymal stem cells[J]. Cell Biol Int. 2015;39(9):1026-1035.
[Crossref] [Google Scholar] [PubMed]
- Lacci KM, Dardik A. Platelet-rich plasma: support for its use in wound healing. The Yale journal of biology and medicine. 2010;83(1):1.
[Google Scholar] [PubMed]
- Miyakoshi N, Kobayashi M, Nozaka K. Effects of intraarticular administration of basic fibroblast growth factor with hyaluronic acid on osteochondral defects of the knee in rabbits. Arch Orthop Trauma Surg.2005;125(10):683-692.
[Crossref] [Google Scholar] [PubMed]
- Akeda K, An HS, Okuma M. Osteoarthritis and cartilage/OARS, Osteoarthritis Research Society. Osteoarthr Cartil. 2006;14(12)):1272-80.
[Google Scholar] [PubMed]
- Lee HR, Park KM, Joung YK, Park KD, Do SH. Platelet-rich plasma loaded hydrogel scaffold enhances chondrogenic differentiation and maturation with up-regulation of CB1 and CB2. J Control Release. 2012;159(3):332-337.
[Crossref] [Google Scholar] [PubMed]
- Siclari A, Mascaro G, Kaps C, Boux E. A 5-year follow-up after cartilage repair in the knee using a platelet-rich plasma-immersed polymer-based implant. Open J. Orthop. 2014;8:346.
[Crossref] [Google Scholar] [PubMed]
- Gope R. The effect of epidermal growth factor & platelet-derived growth factors on wound healing process. Indian J Med Res. 2002;116:201-206.
[Google Scholar] [PubMed]
- Judith R, Nithya M, Rose C, Mandal AB. Application of a PDGF-containing novel gel for cutaneous wound healing. Life sci. 2010;87(1-2):1-8.
[Crossref] [Google Scholar] [PubMed]
- Pujol N, Salle De Chou E, Boisrenoult P, Beaufils P. Platelet-rich plasma for open meniscal repair in young patients: any benefit?. Knee Surg Sports Traumatol. Arthrosc. 2015;23(1):51-58.
[Crossref] [Google Scholar] [PubMed]
- Yokoyama M, Sato M, Tani Y, Yokoyama M, Kokubo M, Yamato M, Okano T, Mochida J. Plateletâ?activated serum might have a therapeutic effect on damaged articular cartilage. J Tissue Eng Regen Med. 2017;11(12):3305-3312.
[Crossref] [Google Scholar] [PubMed]
- Corrado A, Neve A, Cantatore FP. Expression of vascular endothelial growth factor in normal, osteoarthritic and osteoporotic osteoblasts. Clin Exp Med. 2013;13(1):81-84.
[Crossref] [Google Scholar] [PubMed]
- Chim SM, Tickner J, Chow ST, Kuek V, Guo B, Zhang G, et al. Angiogenic factors in bone local environment. Cytokine & growth factor rev. 2013;24(3):297-310.
[Crossref] [Google Scholar] [PubMed]
- Liu Y, Berendsen AD, Jia S, Lotinun S, Baron R, Ferrara N, et al. Intracellular VEGF regulates the balance between osteoblast and adipocyte differentiation. J Clin Investig. 2012;122(9):3101-3013.
[Crossref] [Google Scholar] [PubMed]
- Yang Q, McHugh KP, Patntirapong S, Gu X, Wunderlich L, Hauschka PV. VEGF enhancement of osteoclast survival and bone resorption involves VEGF receptor-2 signaling and β3-integrin. Matrix Biol. 2008;27(7):589-599. [Crossref] [Google Scholar] [PubMed]
- Niida S, Kondo T, Hiratsuka S, Hayashi SI, Amizuka N, Noda T. VEGF receptor 1 signaling is essential for osteoclast development and bone marrow formation in colony-stimulating factor 1-deficient mice. Proc Natl Acad Sci. 2005;102(39):14016-14021. [Crossref] [Google Scholar] [PubMed]
- Mayr-Wohlfart U, Waltenberger J, Hausser H, Kessler S, Günther KP, Dehio C, et al. Vascular endothelial growth factor stimulates chemotactic migration of primary human osteoblasts. Bone. 2002; 30(3):472-477. [Crossref] [Google Scholar] [PubMed]
- Riboh JC, Saltzman BM, Yanke Abdominal Muscle FL, Cole BJ. Result of Leukocyte Focus on the Effectiveness of Platelet-Rich Plasma in the Therapy of Knee Osteo Arthritis. Am J Sports Medication. 2016;44(3):792-800.
- Zhu Y, Yuan M, Meng HY, Wang AY, Guo QY, Wang Y, et al. Basic science and clinical application of platelet-rich plasma for cartilage defects and osteoarthritis: a review. Osteoarthr Cartil. 2013;21(11):1627-1637.
[Crossref] [Google Scholar] [PubMed]
- Mifune Y, Matsumoto T, Takayama K, Ota S, Li H, Meszaros LB, et al. The effect of platelet-rich plasma on the regenerative therapy of muscle derived stem cells for articular cartilage repair. Osteoarthr Cartil. 2013;21(1):175-85.
[Crossref] [Google Scholar] [PubMed]
- Hsieh JL, Shen PC, Shiau AL, Jou IM, Lee CH, Wang CR, et al. Intraarticular gene transfer of thrombospondinâ?1 suppresses the disease progression of experimental osteoarthritis. J Orthop Res. 2010;28(10):1300-1306.
[Crossref] [Google Scholar] [PubMed]
- Zhu Y, Yuan M, Meng HY, Wang AY, Guo QY, Wang Y, et al. Basic science and clinical application of platelet-rich plasma for cartilage defects and osteoarthritis: a review. Osteoarthr Cartil. 2013;21(11):1627-1637.
[Crossref] [Google Scholar] [PubMed]
- Iacob S, Cs-Szabo G. Biglycan regulates the expression of EGF receptors through EGF signaling pathways in human articular chondrocytes. Connect Tissue Res. 2010;51(5):347-358.
[Crossref] [Google Scholar] [PubMed]
- Zhenhai W, Qin F U, Zhonghai Z. Influence of EGF and IGF on articular cartilage cells of rabbit in vitro[J]. China Journal of Orthopaedics and Traumatology. Chin. J. Traumatol. 2003(01):27-29.
- Nonaka K, Shum L, Takahashi I, Takahashi KA, Ikura T, Dashner RA, et al. Convergence of the BMP and EGF signaling pathways on Smad1 in the regulation of chondrogenesis. Int J Dev Biol. 2002;43(8):795-807.
- Huh YH, Kim SH, Kim SJ, Chun JS. Differentiation status-dependent regulation of cyclooxygenase-2 expression and prostaglandin E2 production by epidermal growth factor via mitogen-activated protein kinase in articular chondrocytes. J Biol Chem. 2003;278(11):9691-9697.
[Crossref] [Google Scholar] [PubMed]
- Janssen JN, Batschkus S, Schimmel S, Bode C, Schminke B, Miosge N. The influence of TGF-β3, EGF, and BGN on SOX9 and RUNX2 expression in human chondrogenic progenitor cells. J Histochem Cytochem. 2019;67(2):117-127.
[Crossref] [Google Scholar] [PubMed]
- Smelter E, Hochberg MC. New treatments for osteoarthritis. Curr Opin Rheumatol. 2013;25(3):310-316.
- Pecchi E, Priam S, Gosset M. Induction of nerve growth factor expression and release by mechanical and inflammatory stimuli in chondrocytes: possible involvement in osteoarthritis pain. Arthritis Res Ther. 2014;16(1):16.
[Crossref] [Google Scholar] [PubMed]
- Nico B, Mangieri D, Benagiano V, Crivellato E, Ribatti D. Nerve growth factor as an angiogenic factor. Microvasc Res. 2008;75(2):135-141.
[Crossref] [Google Scholar] [PubMed]
- Dahlin RL, Ni M, Meretoja VV, Kasper FK, Mikos AG. TGF-β3-induced chondrogenesis in co-cultures of chondrocytes and mesenchymal stem cells on biodegradable scaffolds. Biomaterials. 2014;35(1):123-132.
[Crossref] [Google Scholar] [PubMed]
- Walsh DA, McWilliams DF, Turley MJ, Dixon MR, Fransès RE, Mapp PI, et al. Angiogenesis and nerve growth factor at the osteochondral junction in rheumatoid arthritis and osteoarthritis. Rheumatology. 2010;49(10):1852-1861.
[Crossref] [Google Scholar] [PubMed]
- Duan L, Ma B, Liang Y, Chen J, Zhu W, Li M. Cytokine networking of chondrocyte dedifferentiation in vitro and its implications for cell-based cartilage therapy. Am J Transl Res. 2015; 7(2):194.
[Google Scholar] [PubMed]
- Phull AR, Eo SH, Abbas Q, Ahmed M, Kim SJ. Applications of chondrocyte-based cartilage engineering: an overview. BioMed Res Int. 2016.
[Crossref] [Google Scholar] [PubMed]
- KikuchiA Y, Kishida S. Multiplicity of the interactions of Wnt proteins and their receptors. Cell Signal. 2007;19(4):659-671.
[Crossref] [Google Scholar] [PubMed]
- Church V, Nohno T, Linker C, Marcelle C, Francis-West P. Wnt regulation of chondrocyte differentiation. J cell sci. 2002;115(24):4809-4818.
[Crossref] [Google Scholar] [PubMed]
- Hwang SG, Ryu JH, Kim IC, Jho EH, Jung HC, Kim K, et al. Wnt-7a causes loss of differentiated phenotype and inhibits apoptosis of articular chondrocytes via different mechanisms. J Biol Chem. 2004;279(25):26597-26604.
[Crossref] [Google Scholar] [PubMed]
- Hwang SG, Yu SS, Lee SW, Chun JS. Wnt-3a regulates chondrocyte differentiation via c-Jun/AP-1 pathway. FEBS letters. 2005;579(21):4837-4842.
[Crossref] [Google Scholar] [PubMed]
- Day TF, Guo X, Garrett-Beal L, Yang Y. Wnt/β-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Dev Cell. 2005;8(5):739-750.
[Crossref] [Google Scholar] [PubMed]
- Chen Y, Whetstone HC, Youn A, Nadesan P, Chow EC, Lin AC, et al. β-catenin signaling pathway is crucial for bone morphogenetic protein 2 to induce new bone formation. J Biol Chem. 2007;282(1):526-533.
[Crossref] [Google Scholar] [PubMed]
- Iida K, Itoh E, Kim DS. Muscle mechano growth factor is preferentially induced by growth hormone in growth hormone-deficient lit/lit mice. J Physiol. 2004;560:341-349.
[Crossref] [Google Scholar] [PubMed]
- Tong Y, Feng W, Wu Y, Lv H, Jia Y, Jiang D. Mechano-growth factor accelerates the proliferation and osteogenic differentiation of rabbit mesenchymal stem cells through the PI3K/AKT pathway. BMC biochem. 2015;16(1):1-6.
[Crossref] [Google Scholar] [PubMed]
- Luo Z, Jiang L, Xu Y. Mechano growth factor (MGF) and transforming growth factor (TGF)-β3 functionalized silk scaffolds enhance articular hyaline cartilage regeneration in rabbit model. Biomaterials. 2015;52:463-475.
[Crossref] [Google Scholar] [PubMed]
- Deng M, Zhang B, Wang K, Liu F, Xiao H, Zhao J. Mechano growth factor E peptide promotes osteoblasts proliferation and bone-defect healing in rabbits. Int Orthop. 2011;35(7):1099-1106. [Crossref] [Google Scholar] [PubMed]
- Song Y, Xu K, Yu C, Dong L, Chen P, Lv Y, et al. The use of mechano growth factor to prevent cartilage degeneration in knee osteoarthritis. J Tissue Eng Regen Med. 2018;12(3):738-749.
- Skardal A, Zhang J, McCoard L, Xu X, Oottamasathien S, Prestwich GD. Photocrosslinkable hyaluronan-gelatin hydrogels for two-step bioprinting. Tissue Eng Part A. 2010;16(8):2675-2685.
[Crossref] [Google Scholar] [PubMed]
- Han Y, Zeng Q, Li H, Chang J. The calcium silicate/alginate composite: preparation and evaluation of its behavior as bioactive injectable hydrogels. Acta biomaterialia. 2013 Nov 1;9(11):9107-17.
[Crossref] [Google Scholar] [PubMed]
- Gantar A, da Silva LP, Oliveira JM, Marques AP, Correlo VM, Novak S, et al. Nanoparticulate bioactive-glass-reinforced gellan-gum hydrogels for bone-tissue engineering. Mater. Sci. Eng: C. 2014;43:27-36.
[Crossref] [Google Scholar] [PubMed]
- Khanarian NT, Haney NM, Burga RA, Lu HH. A functional agarose-hydroxyapatite scaffold for osteochondral interface regeneration. Biomaterials. 2012;33(21):5247-5258.
[Crossref] [Google Scholar] [PubMed]
- Filardo G, Kon E, Di Martino A. Treatment of knee osteochondritis dissecans with a cell-free biomimetic osteochondral scaffold: Clinical and imaging evaluation at 2-year follow-up. Am J Sports Med. 2013 ;41(8):1786-1793.
- Formica FA, Barreto G, Zenobi-Wong M. Cartilage-targeting dexamethasone prodrugs increase the efficacy of dexamethasone. J Control Release. 2019;295:118-129.
[Crossref] [Google Scholar] [PubMed]
- Nuttelman CR, Tripodi MC, Anseth KS. Dexamethasoneâ?functionalized gels induce osteogenic differentiation of encapsulated hMSCs. Journal of Biomedical Materials Research Part A: An Official Journal of The Society for Biomaterials, The Japanese Society for Biomaterials, and The Australian Society for Biomaterials and the Korean Society for Biomaterials. 2006;76(1):183-195.
[Crossref] [Google Scholar] [PubMed]
- Florine EM, Miller RE, Porter RM, Evans CH, Kurz B, Grodzinsky AJ. Effects of dexamethasone on mesenchymal stromal cell chondrogenesis and aggrecanase activity: Comparison of agarose and self-assembling peptide scaffolds. Cartilage. 2013;4(1):63-74.
[Crossref] [Google Scholar] [PubMed]
- Kim M, Garrity ST, Steinberg DR, Dodge GR, Mauck RL. Role of dexamethasone in the longâ?term functional maturation of MSCâ?laden hyaluronic acid hydrogels for cartilage tissue engineering. J Orthop Res. 2018;36(6):1717-1727.
[Crossref] [Google Scholar] [PubMed]
- Harrell CR, Markovic BS, Fellabaum C, Arsenijevic A, Volarevic V. Mesenchymal stem cell-based therapy of osteoarthritis: Current knowledge and future perspectives. Biomed Pharmacother. 2019;109:2318-2326.
[Crossref] [Google Scholar] [PubMed]
- Sheehy EJ, Mesallati T, Vinardell T, Kelly DJ. Engineering cartilage or endochondral bone: A comparison of different naturally derived hydrogels. Acta biomaterialia. 2015;13:245-253.
[Crossref] [Google Scholar] [PubMed]
Citation: Zhang Y, Cai Z, Chengqiang Y, Wu Y, Ou Y, Wei J (2022) The Role of Growth Factors in Articular Cartilage Repair: A Systematic Review. J Aging Sci. 10:274.
Copyright: © 2022 Zhang Y, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.