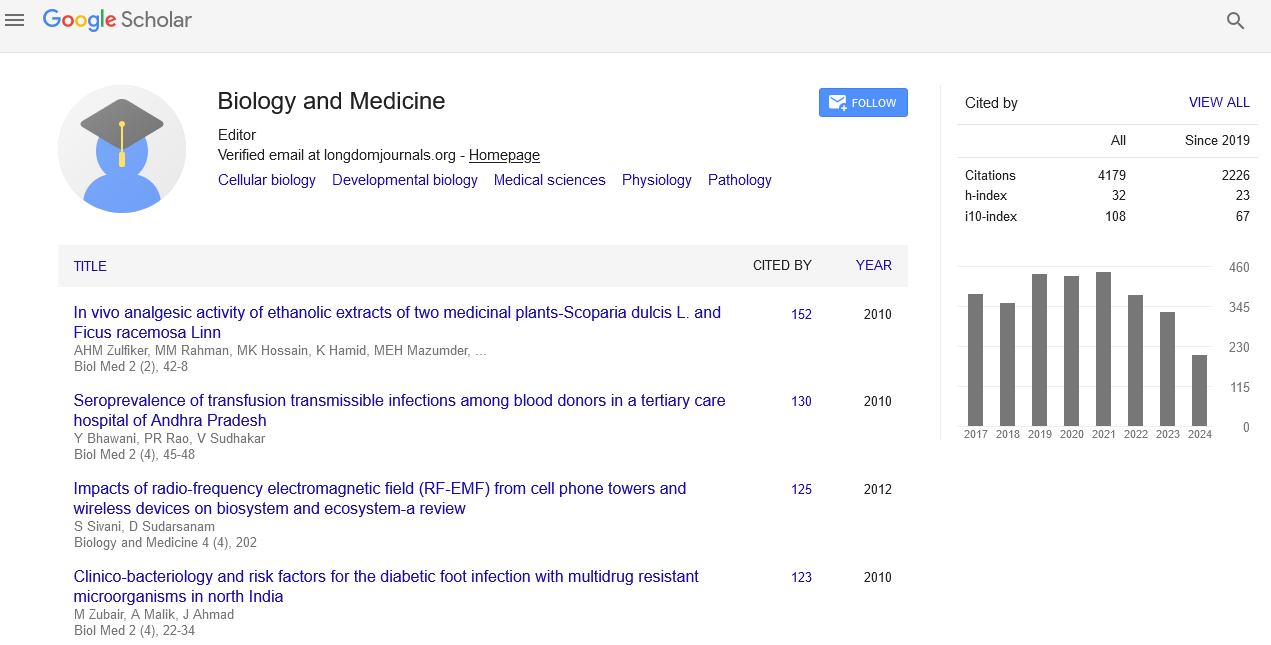

Indexed In
- Open J Gate
- Genamics JournalSeek
- CiteFactor
- Cosmos IF
- Scimago
- Ulrich's Periodicals Directory
- Electronic Journals Library
- RefSeek
- Hamdard University
- EBSCO A-Z
- Directory of Abstract Indexing for Journals
- OCLC- WorldCat
- Proquest Summons
- Scholarsteer
- ROAD
- Virtual Library of Biology (vifabio)
- Publons
- Geneva Foundation for Medical Education and Research
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Research Article - (2023) Volume 15, Issue 8
Enhancing Genetically Engineered Escherichia coli Bioreporters for the Detection of Buried TNT-based Landmines
Eden Amiel*Received: 10-Aug-2023, Manuscript No. BLM-23-22570; Editor assigned: 14-Aug-2023, Pre QC No. BLM-23-22570 (PQ); Reviewed: 28-Aug-2023, QC No. BLM-23-22570; Revised: 07-Sep-2023, Manuscript No. BLM-23-22570 (R); Published: 14-Sep-2023, DOI: 10.35248/0974-8369.23.15.604
Abstract
TNT (Trinitrotoluene) based buried landmines are explosive devices consisting of mainly 2,4,6-Trinitrotoluene (2,4,6-TNT), but also a 2,4-dinitrotoluene (2,4-DNT) impurity. It is reported that some 2,4-DNT impurity vapors slowly leak through land mine covers and into the soil above them, making this compound a potential indicator for the presence of buried landmines. There are several approaches to identify the presence of chemicals in the environment, one of which is based on a microbial detection system. This approach was previously used to address the challenge of remotely identifying buried landmines by applying genetically engineered microbial whole cell biosensors capable of sensing trace signatures of the explosive compound 2,4,6-TNT and its degradation product, 2,4-DNT. Upon exposure to 2,4-DNT, the genetically engineered bacterial reporter strains, containing a genetic fusion between a sensing element for 2,4-DNT (yqjF gene promoter) and a fluorescence/ bioluminescence reporting element (GFP/luxCDABE genes), generate a measurable dose-dependent signal within the whole cell biosensor system. This biological component of a comprehensive remote sensing solution was significantly improved after several rounds of random mutagenesis to the yqjF promoter region using a prone-to-error PCR and selection of the best mutants. In the present project further significant improvements were obtained in signal intensity, response time, and in the limit of detection, following an additional round of random mutagenesis. Two new mutants, named A10 and C5, were identified. The new mutant’s present high induction levels both in terms of Ratio (sample/control) and delta (sample- control), making these strains more amenable to remote detection. To isolate a more sensitive mutant, the 3rd round of random mutagenesis library (about 1000 strains) was re-exposed to lower concentration of 2,4-DNT, but no better candidate was isolated. Possible alternative approaches to lower the detection threshold of the system, such as the use of permeability mutants, are now being tested.
Keywords
Trinitrotoluene; Escherichia coli; Bio reporters
Introduction
Landmines are containers of explosive material with detonating systems that are triggered by contact with a person or vehicle, creating an explosive blast designed to hinder or ultimately immobilize and put the advancement of the enemy into a halt [1]. Mine and countermine technologies and techniques have evolved over the past 3,000 years that by the 20th century, the antipersonnel mine grew into a highly effective weapon and combat multiplier.
History of the weapon
During the First World War the use of battle tanks drove the development of the antitank landmine and later the antipersonnel landmine, designed to prevent enemy soldiers from removing the antitank landmines [2]. Between 1918 and 1939 the development and use of the antipersonnel landmine became a priority among military strategists putting it into use throughout Poland, Russia and Korea. AP’s were then used in World War II to defend military positions and socioeconomic targets, channel the movement of troops to certain routes from which they can’t deviate, and to cause chaos and economic dislocation as political weapons [3]. Subsequently, AP’s were widely deployed around the world in the period from World War II until the treaty banning their use, production, transfer and stockpiling was adopted in 1997 [4]. As a result, dozens of nations have mine-affected land that must be cleared before it can be returned to productive use. Long after wars are over, landmines make land unusable for farming, schools or living, preventing people from rebuilding lives torn apart by conflict [5]. In 2002, the number of landmines planted around the world was estimated to be above 110 million spread in 70 countries, and the time required removing them around 1100 years [6], mainly due to the risks and complications involved in landmines detection and removal.
Explosives trace signatures and detection techniques
Approximately 80% of all landmines are TNT-based, meaning the main explosive found in them is comprised of 2,4,6-Trinitrotoluene (2,4,6-TNT). These also contain additional manufacture impurities and degradations products including 1,3-Dinitrobenzene (1,3- DNB) and 2,4-Dinitrotoluene (2,4-DNT) [7,8]. Leakage occurs through cracks in the mine casing and vapors diffuse through the plastic housing of the mine. Some of the emitted explosive traces will be adsorbed to soil constituents and the rest will travel away from the mine, migrating to the ground surface in vapor form, thus serving as a potential indicator for buried landmines [7,9,10].
The identification of these trace compounds has served as a basis for miscellaneous approaches for landmine biodetection [11]. This work is based on using the environmentally more stable 2,4-DNT as an indicator for TNT-landmines. Landmines detection methods can be divided into two basic classes: Detection of the non- explosive component (mine case or detonator) and detection of the explosive component and its traces. At present, landmine detection is mostly conducted in a manner similar to that employed 60 years ago-based on metal detection and manual characterization of a detected buried object [9,12]. Although its popularity, this method has high false positive rate and it requires immediate proximity to the landmine. Other approaches under development include chemical and physical detection as well as biological detection which are based on the present of trace signatures in the ground above the mine. One of these biological detection techniques is a comprehensive biosensor system.
Biosensors are defined as measurement devices or systems based on the coupling of a biological component with a transducing element, together producing a measurable signal serving as an indicator for the presents of specific chemicals. When compared to analytical methods, biosensors have several advantages including ease of use, low cost, short response times and specificity. All of these combine to allow simplified field applicability [13]. In this study, the biosensor system we used was comprised of whole-cell genetically engineered bacteria assembled by Yagur-Kroll, et al. [14] who’s identified in a previous study two E.coli endogenous gene promoters: yqjF (predicted quinol oxidase subunit) and ybiJ, shown to be responsive to 2,4-DNT. These promoters were selected through the scan of 2,000 E.coli reporter strains library, and genetically fused upstream to either fluorescent or bioluminescence (luxCDABE) reporter genes constructing an E.coli whole-cell sensor strain for 2,4- DNT and 2,4,6-TNT. These were then tested for a dose-dependent response to 2,4-DNT in an aqueous solution, in vapors and in soil. All tests indicated that the endogenous promoters are not sensitive enough for landmine detection in situ, and thus improvement of the sensitivity i.e. lowering the detection threshold is required to consider this method of detection as applicable [14]. When testing the system for a response to explosive traces, selection of suitable reporting genes is required. Bioluminescence allows a much faster and more sensitive detection of the target analyte than fluorescence and thus is easier to work with and more efficient when it comes to lab experiments. The main advantage of fluorescence sensing is reporter stability-once induced, the signal keeps accumulating and may be detected for many hours, in some cases even after the cell death [15,16]. The differences between the two systems derives from the fact that bioluminescence is a measure of enzymatic activity (luciferase enzyme), whereas fluorescence quantifies the presence of a protein (green fluorescent protein) [17]. The kind of results that are achieved with the fluorescence system are better suited for the final, in situ detection system, where sufficient time should allow for the signal (a protein) to accumulate and even improve the sensitivity of the detection system. In the same previous former study [14], the previously constructed E.coli whole-cell sensor strain was significantly improved following three rounds of random mutagenesis to the yqjF promoter region using a prone-to-error PCR (Polymerase Chain Reaction) and selection of the best mutants in a ‘directed evolution’ kind of manner. This approach is based on the creation of new variants by random point mutation, screening of the variants with low levels of 2,4-DNT and selecting the improved variants [18,19]. Following the selection of the improved variants, a dose-dependent experiment is conducted to yet further narrow the search and to select a few of the most enhanced mutants. In this study, further performance improvement of the yqjF promoter was achieved in terms of detection threshold, response time and signal intensity through a 4th round of promoter-directed Random Mutagenesis (RM). Two new mutants, named A10 and C5 were identified, both present high induction levels in terms of Ratio and Delta making these strains more amenable to remote detection.
Each point mutation found in the A10 and C5 strains was than lonely inserted to the promoter region in order to shed some light on the mechanism of action in the induction of yqjF by 2,4-DNT and maybe better understand the possible role of each region in the promoter itself, including possible transcription factors binding sites and other sites important for the transcription of the yqjF gene. Several combinations of numerous point mutations together were also tested for the purpose of learning whether one point mutation can affect the others in a synergetic or an antagonistic kind of way, if at all.
Methods and Materials
Bacterial strains and plasmids construction
The plasmids and bacterial strains employed and their genotypes are listed in Table 1. Below is a description of the construction of the plasmids generated for the purpose of the present study. E.coli K12 strain MG1655 chromosomal DNA was used as a template for promoters amplification in the PCRs. E.coli K12 strain DH5 α was used as a host for the construction of the plasmids and for the random mutagenesis procedure. E.coli K12 strain MG1655 was used as a host for all exposure assays in liquid media (Table 2).
| Strains | Description | Reference |
|---|---|---|
| MG1655 | F-λ- ilvG- rfb-50 rph-1 | Blattner FR et al. [19] |
| DH5α | F- endA1 hsdR17 (rk-,mk+) supE44 thi-1 λ- recA1 gyrA96 relA1 deoR ∆(lacZYA-argF)-U169 ф80dlacZ∆M15 | Grant SG et al. [20] |
Table 1: E.coli strains used and their genotypes.
| Primer | Sequence |
|---|---|
| yqjF-KpnI | 5’ GGTGGTACCCGGTTTTGGCGTATGG 3’ |
| yqjF-SacI | 5’ CGGAGCTCGCCACTCAGGCTGCTGAT 3’ |
Table 2: Primers used in this study.
Random mutagenesis of the yqjF promoter region
Another round of random mutagenesis using error-prone PCR was performed to the improved variant yqjFB2A1#14::lux, already containing several point mutations in its promoter region (Figures S1 and S2), constructed in a previous work following three rounds of random mutagenesis [14]. For the technique to work properly, it was important to use a Taq DNA polymerase which does not have proof-reading ability. Following the error-prone PCR reaction, the PCR products were gel-purified, digested with KpnI and SacI restriction enzymes [18], and ligated into a promoterless pBR2TTS::luxCDABE plasmid (Figure S1), digested with the same restriction enzymes. The products of the ligation reaction were used to transform E.coli strain DH5α to generate the variants library. The library was then screened for improved variants as follows: Each colony was inoculated into an individual well of a 96-well microtiter plate, containing 150 μl of LB medium supplemented with ampicillin (100 μg/ml); two control wells in each plate were occupied by the wild-type strain. This step has been performed by MICROLAB® STAR line (Hamilton, USA) robotic system. The plate was incubated overnight at 37°C with shaking. Next, 10 μl aliquots of the culture were transferred into two opaque white 96- well microtiter plates with a clear bottom: one containing in each well 90 μl of LB supplemented with 25 mg/L 2,4-DNT and the other with 90 μl LB supplemented with 0.16% ethanol. The plates were incubated at 37°C, and optical density and bioluminescence of the cultures were measured every 20 minutes using the MICROLAB robotic system’s plate reader. Fluorescence and luminescence values were recorder and displayed as the instrument’s arbitrary Relative Fluorescence (RFU) and Relative Luminescence Units (RLU), respectively. Blank values were subtracted from the optical density measured, and then the RLU values were divided by the Optical Density (O.D) of each well. RLU (DNT) and RLU (control) denote the luminescence (after division by the O.D) in wells containing bacteria with or without 2,4-DNT, respectively. The induction ratio for each reporter variant was calculated as follows:

The difference in the signal intensity in the presence and absence of the inducer (ΔRLU) was calculated as follows: ΔRLU=RLU(DNT)- RLU(control)
Response sensitivity was determined by calculating the EC200 value, denoting the effective inducer concentration causing a twofold increase in the response ratio. Lower EC200 values reflect greater sensitivity and a lower detection threshold of the tested chemical by the reporter strain employed [20-22]. Reporter variants that displayed a higher induction ratio, a lower 2,4-DNT detection threshold, or a shorter response time than the wild type were selected for further analysis by exposure to a 2,4-DNT concentration series. The selection procedure of the best variants for further testing was done using an algorithm in MATLAB, as explained below (Figure S2).
Genetic algorithm as a part of programmed evolutionary solution
A Genetic Algorithm (GA) script containing multiple functions was developed based on the previous work of Moskovitz and Elad. The previously programmed algorithm had some limitations that this current GA was able to resolve (‘Matlab’ in supplements). The improved GA served us as a search method that mimics the process of directed biological selection. The algorithm operates on a population of potential variants applying the principle of ‘survival of the fittest’ to find better and better candidates suited for our needs. In our fitness-based selection model, the fitness value actually represents the mutant’s ability to detect explosive trace signatures rather than its traditional definition used in several other models [23]. For this, a customized function that assigns fitness values for each variant was developed. At first, the script automatically calculates the induction ratio and the ΔRLU for the first 180 minutes of measurements and creates two kinetic graphs for each one of the variants: one for the ratio and one for the ΔRLU (Figures S3-S6). Each graph is represented by a function equation -f(x). Next, the fitness values/ranking (FitnV) of each variant is calculated by the equation:

That is the sum of the differences between adjacent elements of X along the function f(x). This value is basically a number that describes how much the RLU per OD changed over time and is a good estimation for the mutant’s ability to detect landmines. For fitness-based competition, the FitnV values were organized into a list in descending order. The first 10 values of the two lists (ratio and ΔRLU) were intersected, and the variants with the highest FitnV value in both lists were selected for further analysis.
Exposure to a 2,4-DNT concentration series
The bacterial reporter strains were grown overnight at 37°C in LB medium with 100 μg/ml ampicillin. Then, the cells were diluted 100-fold in fresh medium, and regrown with shaking at 37°C to an early exponential growth phase (O.D600≈0.2). In the next step an opaque white 96-well microtiter plates have been filed, so that each well contained 50 μl of a predetermined concentration of 2,4-DNT dissolved in 200 mM Tris-base pH 7 and 5 μl of 10 concentrated LB media. Into each well, culture aliquots (45 μl) were transferred. Luminescence was measured at 37°C every 10 minutes using a VICTOR2 plate reader (Wallac, Turku, Finland). All experiments were carried out in duplicate, and were repeated at least 3 times. To assess the activity of the tested reporter strains, ΔRLU and induction ratio were calculated as explained above.
Single and multiple-point mutations
For the study of the effect of specific single nucleotide alterations, primers harboring the Point Mutations (PM) of interest were used to perform a standard PCR, inserting PM’s to specific areas in the process. All insertions were confirmed by sequencing.
Results
It was shown in a previous study that sensor performance measured by signal intensity, time of detection, and/or detection threshold, could be improved by a directed evolution procedure based on the insertion of random mutations in the promoter element of the sensor [18]. This procedure has been applied to the yqjF promoter prior to this study, performing three rounds of Random Mutagenesis (RM) PCR and ultimately generating an enhanced sensor called yqjFB2A1#14::lux [14]. In the current study, a 4th round of RM was performed using yqjFB2A1#14::lux as the template for the error-prone PCR procedure, in order to further improve this sensor’s performance. Following the procedure, several candidates were chosen for an exposure test to a 2,4- DNT concentration series. The best performers obtained from the 4th round RM followed by an exposure to a 2,4-DNT concentration series were the variants yqjF4th-A10::lux and yqjF4th-C5::lux (Figure 1).

Figure 1: Induction of the original yqjFwt and improved variants
by 2,4-DNT: Dose-dependent response. A) increase in luminescence
intensity (ΔRLU); b) Response ratio. The data presented are maximal
values obtained in the course of a 300minutes exposure. Note: ( ):
MG/Fwt::lux, (
):
MG/Fwt::lux, ( ): MG/FB2A::lux#14, (
): MG/FB2A::lux#14, ( ): MG/F124-133:lux, (
): MG/F124-133:lux, ( ):
MG/F124-138:lux, (
):
MG/F124-138:lux, ( ): MG/F213::lux, (
): MG/F213::lux, ( ): MG/F2138:lux.
): MG/F2138:lux.
For A10, the response ratio value was up to 42-fold higher than the wild type (287 as compared to 6.8 for 50 mg/L 2,4-DNT), the ΔRLU values were over 400-fold higher (1,167,694 as compared to 2,795 for 50 mg/L 2,4-DNT). For C5, the response ratio value was up to 62-fold higher than the wild type (421 as compared to 6.8 for 50 mg/L 2,4-DNT), the ΔRLU values were over Furthermore, the response time for 25 mg/L 2,4-DNT (Figures 2 and 3), defined as the time the response ratio reaches a value of 2 in this concentration, was shortened from about 100 minutes in the WT to approximately 70 minutes in both A10 and C5, and as opposed to 80 minutes in the former yqjFB2A1#14 mutant (Table S1). The response sensitivity/detection threshold indicated by the EC200 was considerably improved as well, with EC200 values reaching as low as 4.1 mg/L of 2,4-DNT (Figure 2). Sequencing and comparing the promoter regions of the two improved variants revealed that yqjF4th-A10::lux has an addition of one point mutation compared to the former yqjFB2A1#14::lux and yqjF4th-A10::lux has four additional ones (Figure S2). Using different bioinformatics data bases such as RegulonDB [24] and Ecocyc it was shown in a previous study, that the promoter yqjF contains three predicted transcription units, two of which bind the RNA polymerase sigma 32 factor (yqjFp3 and yqjFp4) and one binds RNA polymerase sigma 70 factor (yqjFp2) [14,25]. Of the new muta-tions originated in this study, only one found in C5 was located in a predicted regulatory element of the -10 region of σ32-dependent yqjFp4.

Figure 2: Detection threshold, expressed as EC200-the sample concentration yielding a ratio of 2 (measured over 120 minutes).

Figure 3: Response ratio of the original yqjFwt and improved variants
in liquid phase over 300 minutes, calculated at 25 mg/L of 2,4-DNT. Note: ( ): WT, (
): WT, ( ): First round of evolution, (
): First round of evolution, ( ): Second round
of evolution, (
): Second round
of evolution, ( ): Third round of evolution, (
): Third round of evolution, ( ): Fourth round of
evolution- A10, (
): Fourth round of
evolution- A10, ( ): Fourth round of evolution-C5.
): Fourth round of evolution-C5.
Point mutations-Individual and multiple effects
To better understand the effects of each nucleotide alteration that was made in each round of the RM process and for the purpose of using this knowledge to perceptively engineer an even more enhanced sensor, individual and multiple combinations effects of all point mutations were tested.
The dose-dependent max response ratio over 240 minutes of all 14 point mutations that were inserted and tested individually can be seen in Figure S3, while the response ratio at 100 mg/L 2,4-DNT of a selected few is also presented (Figure 4). In the same sense, the dose-dependent max ΔRLU over 240 minutes of all combinations made between numbers of PMs is shown in Figure S3. The data of a selected few necessary for the discussion are presented as well (Figure 5). Although the ΔRLU of the mutant MG/F124-F133 reached peaked high values of 4 × 106, it presented a very low response ratio of about 2, possibly due to its high background values -250 times higher than that of the WT (Figure S4). In all tests the mutant MG/F138 lowered both the ΔRLU and the response ratio values, but when calculating the EC200 it showed an increase capability regarding the sensitivity of the sensor (Figure 6).

Figure 4: Response ratio of the original yqjFwt and selected individual point mutations depict from various strains in liquid phase calculated at a concentration of 100 mg/L 2,4-DNT. Data is max value over 240 minutes.

Figure 5: Increase in luminescence intensity (ΔRLU). Dose-depndent
response of the original yqjFwt and selected individual point mutations
and their combinations depicted from various strains in liquid phase,
maximal values obtained over 240 minutes. Note: (): MG/Fwt::lux,
(): MG/FB2A::lux#14, (): MG/F124-133:lux, (): MG/F124-
138:lux, (): MG/F213::lux, (): MG/F2138:lux.

Figure 6: Detection threshold, expressed as EC200-the sample concentration yielding a ratio of 2 (measured over 120 minutes). The original yqjFwt and selected combinations of several individual point mutations taken from different strains in liquid phase.
Discussion
Directed evolution by means of random mutagenesis was conducted in this study as one of several solutions proposed in a former study by Yagur-Kroll [14], to possibly further enhance the sensor’s performance for the detection of buried TNT-based land mines. Despite the fact that throughout the course of this study we focused our efforts on the E.coli reporter strain harboring the pBR2TTS:yqjF::luxCDABE plasmid, transformation of the detection system from luminescence luxCDABEbased to a fluorescence GFP-based for the purpose of field testing was performed and experimented in the lab as well. In addition, to isolate a more sensitive mutant the 3rd round of random mutagenesis library (about 1000 strains) was re-exposed to lower concentration of 2,4-DNT and scanned with the improved algorithm we have developed, but no better candidate was found. From the 4th round of RM two new mutants-A10 and C5, were identified; both present high induction levels in terms of Ratio (sample/control) and delta (sample-control). Further significant improvements were obtained in signal intensity, response time and the limit of detection as well, making these strains more amenable to remote detection of land mines. However, the limits of detection described were still above the extremely low vapor concentrations (in the parts per trillion range) of the trace compounds reported to exist above buried land mines [9,26] (Figure 2).
When comparing the max ΔRLU received over 300 minutes to the corresponding max response ratio it can be observed (Figure 1) that the two new mutants are superior to the last yqjFB2A1#14 mutant in terms of ratio but not in terms of delta. It can be assumed that this result is not necessarily bad due to mutant’s #14 high background values (Figure S4) which, in turn, contributes to its relatively high delta values and low ratio values (Figure 3). This suggests that by lowering the background values while keeping the induction signal intensity high as seen in the new mutants, it might be easier to detect the trace signatures found above buried landmines in the field, due to a higher response ratio. Of the four new PMs inserted by the 4th RM in the C5 strain, one PM named F3 was located in a predicted regulatory element of the prinbow box (-10 region) of σ32-dependent yqjFp4 (Figure S2). When tested the individual effect of this mutant it showed a decreased response than that of the WT in both terms of ΔRLU and EC200. Three other PMs (F124, F128, F133) displaying promising results, along with others, were introduced in the former 3 rounds of RM done prior to this work [14]. These mutants were later found to be located in the -35 region of the σ70-dependent yqjFp2 transcription unit. In the current study, each of these three PMs was tested individually, every single one presenting a vast enhancement in its response by all aspects. Other results from the individual and multiple point mutations tests suggests that although mutant F138 had a negative effect on the performance of the sensor in terms of delta and ratio it seemed to have increased the sensitivity of the sensor when combined with the promising F124-F133 mutant, consisting 3 PMs in the -35 region as explained above (Figures 4-6). This result is the possible outcome of the reduction in the background values displayed by the F138 mutant (Figure S4) and can serve as a basis for a new approach to further increase the sensor’s sensitivity- by lowering the background values of the strain. In addition, it was surprising to see that the only PM inserted in the A10 strain (F49) had little or no effect when inserted individually (Figure 4), but when combined with the former PMs found in yqjFB2A1#14 mutant it had an enhancing response to the former one. This suggests that a specific set of multiple mutations can have a synergetic affect than that of each one individually.
It was concluded in an earlier study conducted by Moskovitz [27] that due to the dramatically improved sensor’s response results achieved when mutagenesis was applied to the -35 region of the σ70- dependent yqjFp2 transcription unit, sigma 70 plays a major role in the transcription of this gene and that the possible outcome of the new PMs introduced in mutant F124-F133 could be an increment in the affinity of the RNA polymerase-σ70 holoenzyme to the yqjF promoter. However, based on the results acquired in this study I suggest that a different possible mechanism of action for the induction of the yqjF promoter by 2,4-DNT takes place. It might be possible that the induction of this promoter occurs through the σ32-dependent yqjFp4 transcription unit and not the σ70-dependent yqjFp2 transcription unit as proposed before. This conclusion is based on the following observations and assumptions: (1) The F3 mutant which introduced a nucleotide alteration in the prinbow box (-10 region) of σ32-dependent yqjFp4 resulted in a decreased activity of the promoter when compared to the WT. (2) Sigma factor subunit binds transiently to the core RNA polymerase enzyme and directs it to specific binding sites. Because specific sigma factors are needed at specific promoters, the use of sigma factors gives E.coli a powerful mechanism to regulate a group of genes involved in a specific function, such as the Heat Shock (HS) response genes. RNA polymerase binds to the promoters of these genes only when σ70 is replaced with the σ32 subunit (specific for heat shock promoters). Which different σ subunits will bind the core enzyme is determined by several factors including its relative cellular levels compared to other σ subunits levels [29]. During normal exponential growth, σ70 accounts for 60%-95% of the total pool of cellular sigma factors with approximately 700 molecules per cell while σ32 accounts for less than 10 molecules per cell [28,29]. In vitro experiments at low temperatures have shown that σ32 and σ70 have similar affinities for RNA polymerase-core binding [30]. This fact suggests that σ32 competes with σ70 for RNA polymerase binding and thus lowering the levels of free σ70 will provide σ 32 acompetitive advantage. To my understanding, this can be the case in our research in which by making mutation in the -35 region of the σ70-dependent transcript we’re not increasing the σ70-RNAP holoenzyme’s affinity to the promoter but rather decreasing its affinity. What this does is that it leaves the σ70 subunit bound to the core RNA polymerase rendering it unavailable and not allowing it to dissociate, as initiation of transcription is required for the release of the sigma factor. Consequently, the cellular levels of free σ70 should drop, increasing the relative cellular levels of σ32. This should initiate the transcription of σ32-dependent transcripts, as explained above. In this way, the results we observed could be the result of down-regulating a specific transcript, which in turn up-regulates the transcription of a different, possibly HS related transcript. This explanation can also give us an answer as to why an endogenous E.coli promoter is strongly and specifically induced by synthetic chemicals that are not usually found in its environment. A tolerance mechanism against nitrogen compounds and exploitation of the nitro groups as a nitrogen source or the aromatic ring as a carbon source were previously suggested [1]. It can now be assumed that this is true in the case of cell’s exposure to various environmental insults, such as nitrogen or carbon starvation which can induce σ32 [31] that will subsequently bind RNAP and initiate the transcription of the yqjF gene. The ultimate response of the cell for this case is scavenging: when a particular nutrient becomes limiting, E.coli increases the production of proteins that forage for the limiting nutrient [32,33]. One of these scavenging proteins might include the yqjF gene which translates to an inner membrane protein with four predicted transmembrane domains. In addition, it was shown that E.coli is able to use 2,4,6-TNT as the sole nitrogen source, making this proposed explanation more probable. Although this might be the case, further research is clearly required in order to understand the role of yqjF in the metabolism of TNT and 2,4-DNT. Determination of the relative cellular levels of σ32 by means of Western blotting should be considered as a method to further research or to disprove this hypothesis.
Conclusion
In conclusion, by applying the directed evolution approach, further improvements were made towards an operative TNT-based landmine detection system. The new variants displayed higher signal intensity, responded faster and were able to detect lower 2,4-DNT concentrations. Though not yet sufficiently sensitive to detect the low levels reported to exist above buried land mines, the efficiency of the directed evolution technique on the promoter region demonstrated in this work is expected to get us closer to a fully functioning landmine detection system.
Future research endeavors should focus on increasing the sensitivity of the system, possibly by the use of mutant strains of several membrane proteins as a host for the genetic fusion to ultimately lower the efflux of 2,4-DNT from the cell. Another approach is to increase the influx levels of the substance through the fusion of the integral membrane protein OmpF porin gene to an IPTG-inducible lacZ gene promoter. Moreover, to further test the effects of multiple mutations individually inserted, a better understanding of the possible role each mutant acquires is necessary due to the high possible combinations that can be made and tested  .
.
References
- Lalush C. Whole-Cell TNT Landmine Bioreporter: Characterization and Improvement. Hebrew University of Jerusalem. 2012.
- Pearn J. The devil's gardens: A history of landmines. BMJ. 2002;324(7353):1589.
- Croll, Mike. The history of landmines. Pen & Sword, 1998.
- Anderson K. The Ottawa convention banning landmines, the role of international non-governmental organizations and the idea of international civil society. Eur J Int Law. 2000;11(1):91-120.
- Strobel, Warren. “Land Mines, Cheap and Deadly, Wreak Terror in the Third World.” Insight on the News. 1994: 10(39).
- Gao P, Collins LM. A two-dimensional generalized likelihood ratio test for land mine and small unexploded ordnance detection. Signal Process. 2000;80(8):1669-1686.
- Sylvia, James M., et al. “Surface-enhanced Raman detection of 2,4-dinitrotoluene impurity vapor as a marker to locate landmines.” Anal Chem. 2000;5834-5840.
[Crossref] [Google Scholar] [PubMed]
- Göth, Ann, Ian G. McLean, and James Trevelyan. “How do dogs detect landmines.” 2003;195-208.
- MacDonald J, Lockwood JR, McFee J, Altshuler T, Broach T, Carin L, et al. Alternatives for landmine detection. Santa Monica, CA: Rand. 2003.
- Phelan JM. Chemical sensing for buried landmines-fundamental processes influencing trace chemical detection. Sandia National Lab.(SNL-NM), Albuquerque, NM (United States); Sandia National Lab (SNL-CA), Livermore, CA (United States). 2002.
- Habib MK. Controlled biological and biomimetic systems for landmine detection. Biosens Bioelectron. 2007;23(1):1-8.
[Crossref] [Google Scholar] [PubMed]
- Harmon RS, DeLucia FC, LaPointe A, Winkel RJ, Miziolek AW. LIBS for landmine detection and discrimination. Anal Bioanal Chem. 2006;385:1140-1148.
[Crossref] [Google Scholar] [PubMed]
- Dennison MJ, Turner APF. “Biosensors for environmental monitoring". Biotechnol Adv. 1995;13(1):1-12.
- Yagur-Kroll S, Lalush C, Rosen R, Bachar N, Moskovitz Y, Belkin S. Escherichia coli bioreporters for the detection of 2, 4-dinitrotoluene and 2, 4, 6-trinitrotoluene. Appl Microbiol Biotechnol.2014;98:885-895.
[Crossref] [Google Scholar] [PubMed]
- Sagi E, Hever N, Rosen R, Bartolome AJ, Premkumar JR, Ulber R, et al. Fluorescence and bioluminescence reporter functions in genetically modified bacterial sensor strains. Sens Actuators B: Chem. 2003;90(3):2-8.
- Hakkila K, Maksimow M, Karp M, Virta M. Reporter genes lucFF, luxCDABE, gfp, and dsred have different characteristics in whole-cell bacterial sensors. Anal Biochem. 2002;301(2):235-242.
[Crossref] [Google Scholar] [PubMed]
- Belkin S. Microbial whole-cell sensing systems of environmental pollutants. Curr Opin Microbiol. 2003;6(3):206-212.
[Crossref] [Google Scholar] [PubMed]
- Yagur‐Kroll S, Bilic B, Belkin S. Strategies for enhancing bioluminescent bacterial sensor performance by promoter region manipulation. Microb Biotechnol. 2010;3(3):300-310.
[Crossref] [Google Scholar] [PubMed]
- Blattner FR, Plunkett III G, Bloch CA, Perna NT, Burland V, Riley M, et al. The complete genome sequence of Escherichia coli K-12. Science. 1997;277(5331):1453-1462.
[Crossref] [Google Scholar] [PubMed]
- Grant SG, Jessee J, Bloom FR, Hanahan D. Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants. Proc Natl Acad Sci. 1990;87:4645-4649.
[Crossref] [Google Scholar] [PubMed]
- Belkin S. A panel of stress-responsive luminous bacteria for monitoring wastewater toxicity. Bioluminescence methods and protocols. 1998:247-258.
[Crossref] [Google Scholar] [PubMed]
- Belkin S, Smulski DR, Dadon S, Vollmer AC, Van Dyk TK, Larossa RA. A panel of stress-responsive luminous bacteria for the detection of selected classes of toxicants. Water Research. 1997;31(12):3009-3016.
- Fonseca CM. Multiobjective Genetic Algorithms with Application to Control Engineering Problems. 1995.
- Salgado H, Peralta-Gil M, Gama-Castro S, Santos-Zavaleta A, Muniz-Rascado L, García-Sotelo JS, et al. Omics data sets, evolutionary conservation, regulatory phrases, cross-validated gold standards and more. Nucleic acids Res. 2013;41:203-213.
[Crossref] [Google Scholar] [PubMed]
- Keseler IM, Mackie A, Peralta-Gil M, Santos-Zavaleta A, Gama-Castro S, Bonavides-Martínez C, et al. EcoCyc: Fusing model organism databases with systems biology. Nucleic Acids Res. 2013;41:605-612.
[Crossref] [Google Scholar] [PubMed]
- Leggett DC. Release of explosive-related vapors from land mines. US Army Corps of Engineers, Engineer Research and Development Center. 2001.
- Moskovitz Y. Improving Escherichia coli bioreporters for the detection of landmines. Hebrew University of Jerusalem. 2013.
- Jishage M, Iwata A, Ueda S, Ishihama A. Regulation of RNA polymerase sigma subunit synthesis in Escherichia coli: intracellular levels of four species of sigma subunit under various growth conditions. J Bacteriol. 1996;178(18):5447-5451.
[Crossref] [Google Scholar] [PubMed]
- Nelson DL, Lehninger AL, Cox MM. Lehninger principles of biochemistry. Macmillan. 2008.
- Arsène F, Tomoyasu T, Bukau B. The heat shock response of Escherichia coli. Int J Food Microbiol. 2000;55(3):3-9.
- Jenkins DE, Auger EA, Matin A. Role of RpoH, a heat shock regulator protein, in Escherichia coli carbon starvation protein synthesis and survival. J Bacteriol. 1991;173(6):1992-1996.
[Crossref] [Google Scholar] [PubMed]
- Yin H, Wood TK, Smets BF. Reductive transformation of TNT by Escherichia coli: pathway description. Appl Microbiol Biotechnol. 2005;67:397-404.
[Crossref] [Google Scholar] [PubMed]
- Peterson CN, Mandel MJ, Silhavy TJ. Escherichia coli starvation diets: Essential nutrients weigh in distinctly. J Bacteriol. 2005;187(22):7549-7553.
[Crossref] [Google Scholar] [PubMed]
Citation: Amiel E (2023) Enhancing Genetically Engineered Escherichia coli Bioreporters for the Detection of Buried TNT-based Landmines. Bio Med. 15:604.
Copyright: © 2023 Amiel E. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.