
Indexed In
- Academic Journals Database
- Open J Gate
- Genamics JournalSeek
- JournalTOCs
- China National Knowledge Infrastructure (CNKI)
- Scimago
- Ulrich's Periodicals Directory
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- MIAR
- University Grants Commission
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
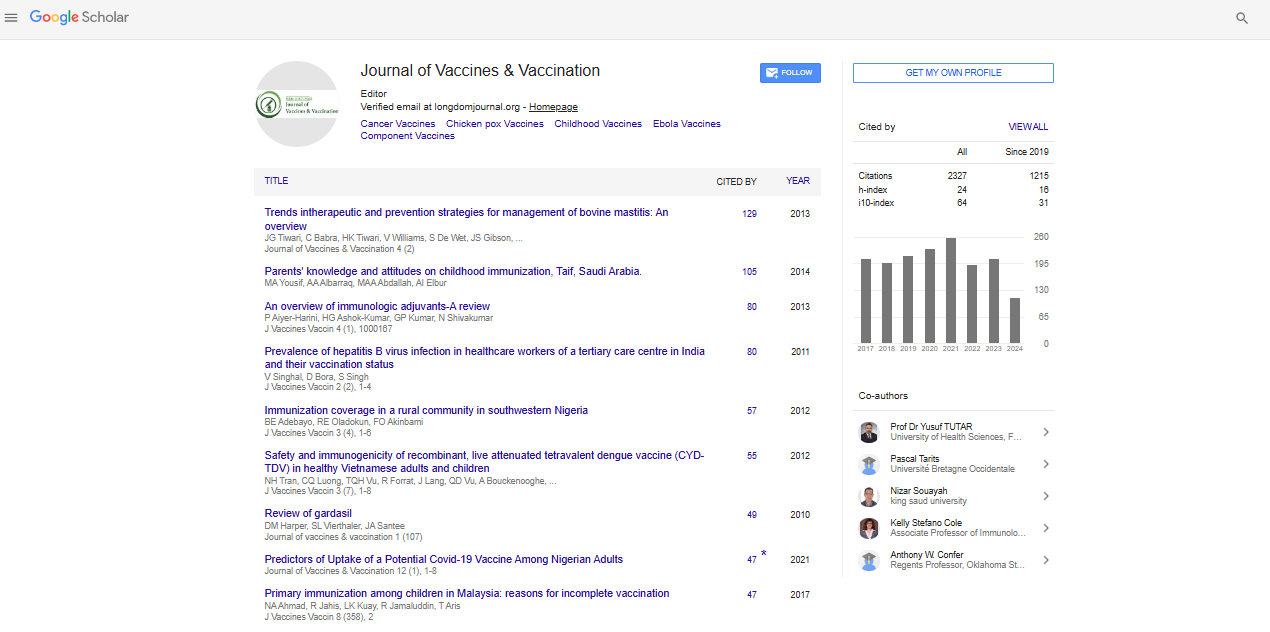
Research Article - (2022) Volume 13, Issue 4
Recent Advances for Vaccine Production in Plants Using Geminiviruses
Maedeh lotfipour*Received: 06-Jun-2022, Manuscript No. JVV-22-001-PreQc-22; Editor assigned: 08-Jun-2022, Pre QC No. JVV-22-001-PreQc-22 (PQ); Reviewed: 23-Jun-2022, QC No. JVV-22-001-PreQc-22; Revised: 30-Jun-2022, Manuscript No. JVV-22-001-PreQc-22 (R); Published: 07-Jul-2022
Abstract
Plants are utilized as a cheap, safe, and efficacious production platform for vaccines and other therapeutic proteins in lowincome countries that do not have access to modern medicine. A simple way to rapidly produce high-level pharmaceutical proteins in plants is to use plant virus expression vectors because their genomes are small and easily manipulated. This study discusses several plant expression systems based on geminiviruses that have been widely developed for vaccine generation and other industrial proteins, as well as recent advances in this field are also presented. Most plant viruses that have been engineered as expression vectors to generate vaccines and other pharmaceutical proteins have RNA-positive genomes, and DNA-based viral vectors are less commonly used because their genomes have a restricted capacity to carry foreign genes. However, the developments of deconstructed vectors have recently led to rapid advances using plant DNA viruses for protein expression. This study discusses the latest progressions in geminiviral expression systems and their use in the production of pharmaceutical proteins.
Keywords
Plant DNA virus; viral vector; Vaccine; Plant
Abbrevations
mAbs: Monoclonal Antibodies; ORF: Open Reading Frame; GOI: Gene Of Interest; AlcR: Alcohol Receptor Gene; LIR: Long Intergenic Region; SIR: Short Intergenic Region; LB: Left Border; RB: Right Border; UTRs: Untranslated Regions; ET: Extension Terminator
Introduction
Plant expression systems used to produce drugs, vaccines, monoclonal Antibodies (mAbs), and other therapeutic proteins have progressed over the past 20 years and become a serious competitor to other eukaryotic systems based on yeast, bacteria, insect, or mammalian cells [1]. Plant systems make it possible to produce large volumes of pharmaceutical proteins in the shortest time and in a completely safe, efficient, and low-cost way. Plant cells are easy to culture in vitro without the need for expensive equipment and controlled environmental conditions. Their growing media are composed of simple nutrients such as sugar and minerals and do not require human- or animal-derived components, thus reducing the risk of contamination with their associated pathogens [2]. Because the secretory pathway and endomembrane system in plant cells is like that of mammalian cells, they can perform suitable posttranslational modifications of recombinant proteins such as glycosylation and correct processing, which are crucial for their stability and function [3]. Plant-derived vaccines for humans and animals can be kept at room temperature without loss of activity, and some of them produced in cereal seeds remain stable over long storage times. These features facilitate the large stockpiling of vaccines against pandemic infectious diseases worldwide and, provide an opportunity to make plant biologics for the developing countries that need access to cheaper medicines [4].
Vaccine candidates or therapeutic proteins were first generated by using a stable transgenic expression system. This approach involved the integration of the target genes into the nuclear or chloroplast genomes, resulting in a high expression level of recombinant proteins in transgenic plants. Although genetically modified plants can supply a permanent source of vaccines, the long process required to generate transgenic lines and concerns about the effects of transgene escapes on the natural environment has led to the use of another technology called transient expression systems based on plant viral vectors [5]. These viral vectors were designed in different ways, from which two types of strategies have been developed so far. The first strategy exploits the unmodified full-length virus that maintains all the genes essential for infection and systemic virulence in its hosts; the gene encoding the protein or peptide of the target is expressed either as part of a fusion protein with a viral protein or alone through an extra sub-genomic promoter integrated into the viral genome. It is called the full-virus strategy or first-generation plant viral expression vectors. This technology can be limited because of the foreign protein size and biosafety concerns because autonomous viral vectors may spread to other plants. The next strategy is second-generation viral vectors or deconstructed virus strategy, in which vectors are designed by replacing the foreign gene of interest with an Open Reading Frame (ORF) of the genome virus that is unnecessary for virus replication. In this method, only the viral elements required for the successful expression of the desired gene are carried in the vector structure, and other needed functions such as DNA delivery are provided by non-viral elements [6]. Recombinant gene is introduced into a high percentage of cells through agro infiltration of deconstructed expression vectors using a syringe into host leaves, or vacuum infiltration into whole plants, resulting in high protein production within a few days. This strategy can overcome the limitations of the classical expression system. Viruses possess small genomes that are more comfortable to manipulate than plant genomes, and plant infection with modified viral vectors is easier and faster than regenerating stable transgenic lines. In addition, deletion of genes encoding movement and/or capsid proteins from the virus constructions reduces foreign proteins’ transmission possibility into the wild populations [7].
Methodology
Geminivirus structure and replication
are a family of plant viruses with genomes comprising one (monopartite) or two (bipartite) single-stranded (ss) circular DNAs of 2.5-5.2 kb, which are encapsidated within twinned semi-icosahedral particles. They are often associated with circular ssDNA molecules of about 1.3 kb, namely alpha satellite and beta satellites that depend on helper viruses for replication, movement, and packaging [8]. All geminivirus genome has a conserved Intergenic Region (IR) containing a stem-loop structure within the origin of replication (ori), as well as promoter elements responsible for gene transcription in both the virion-sense and complementary-sense strands. The coding capacity of geminiviral genomes is limited, ranging from 4 to 8 viral proteins encoded by partially overlapping Open Reading Frames (ORFs) (Figure 1). The complementary sense-strand encodes the replication-associated protein (Rep, ORF C1 or ORFs C1/C2 with spliced intron) and the replication-associated protein A (Rep A, ORF C1), which are both required for viral DNA replication, C2 protein that acts as a transcriptional activator protein (TrAP, ORF C2), a replication enhancer protein (REn, ORF C3), C4 protein, which functions as a pathogenicity determinant protein (ORF C4) and C5 protein (ORF C5). The virion-sense strand contains three ORFs, V1 (AV1), which codes for the Coat Protein (CP); V2 (AV2) is involved in virus movement (MP) and V3 acts as an inhibitor of gene silencing [9,10].

Figure 1: Genome organization of genus Mastrevirus (family Geminiviridae) is illustrated as circular (a) and Linear (b).
Geminiviruses are replicated in the nucleus by the plant’s DNA synthesis machinery. Their replication cycles involve the conversion of ssDNA into dsDNA replicated form by host enzymes and the replicative form acts as a template for viral transcription and a template for amplifying ssDNA genomes by a Rolling Circle Replication (RCR) mechanism. Rep is the only essential viral factor required for replication and initiates ssDNA synthesis in the RCR stage by introducing a nick in the origin viral strand replication between nucleotides 7 and 8 within the nanomer motif (with the sequence TAATATTAC). After cleavage, the Rep protein remains bound to the 51 phosphate end of the nicked strand, and, thus creating a free 31OH end that serves as a primer for the synthesis of nascent viral strands by host DNA polymerase. Rep also has a joining activity that it functions as a terminus to liberate the displaced viral strand and ligates it to make the circular form [11,12]. The RCR mechanism can produce very high viral genome copy numbers (up to 100 copies/cell), making geminiviruses a robust and efficient vector for expressing high levels of foreign therapeutic proteins.
Open Reading Frames (ORFs) on the Virion-sense (V) or Complementary-sense (C) strand and their respective protein products are displayed by color. V1, Movement Protein (MP); V2, Coat Protein (CP); C1, Replication-associated protein A (RepA); C1/C2 with intron splicing, Replication-associated protein (Rep). The Long Intergenic Region (LIR) contains the origin of replication and the bidirectional promoter elements required to initiate gene transcription from both strands. The Short Intergenic Region (SIR) is the origin of C-strand replication and has polyadenylation signals to terminate transcription [13].
Vectors based on geminiviruses
Geminivirus replicon vectors have been exploited mostly to amplify target gene copy numbers in plant cells including Bean yellow dwarf virus, Beet curly top virus, Maize streak virus, Wheat dwarf virus, and Tobacco yellow dwarf virus.
Bean yellow dwarf virus
Bean Yellow Dwarf Virus (BeYDV) is a successful candidate for expression systems based on plant viral vectors and BeYDV-derived constructs have been continuously manipulated during research to achieve the high performance of recombinant proteins. In the field of “vaccine proteins, initial studies involved in the design of the p35SRep/RepA expression cassette containing Large Intergenic Region (LIR), Short Intergenic Region (SIR), and intact Rep gene under CaMV 35S promoter control, as well as pBYSEB cassette including a SEB expression cassette, resides between two copies of the LIR and one copy of the SIR in the form “LIRSEB- SIR-LIR” (knows as LSL vector). SEB is a synthetic version of the Staphylococcus Endotoxin B gene that causes toxic shock syndrome (TSS) in humans. Co-bombarded of tobacco NT-1 cells with these cassettes together resulted in a higher level of SEB expression (~20-fold) than NT-1 cells bombarded with pBYSEB alone [14]. Similarly, Rep of BeYDV can enhance the expression of Norwalk Virus Capsid Protein (NVCP) from the LSL cassette in NT-1 cells and also potato plants when supply from a separate expression cassette tightly controlled by the alcohol inducible alcA promoter. Using this promoter, the induction of Rep expression can be manipulated in such a way that can provide the target proteins to accumulate only in the specific plant tissue and prevents toxicity issues in transgenic plants, as well as escape gene silencing which occurs because of an increase in episomal copy number [15-17]. Gene silencing is a limiting factor for the application of geminiviral vectors in expression systems because it can reduce the level of RNA transcript and thus negatively affect the system’s performance. To solve this problem, a silencing suppressor of plant viruses has been incorporated into the expression systems to avoid the detrimental effects of silencing. For example, Huang et al. 2009 developed a three-vector replicon system with high-yield production of recombinant virus-like particles (VLPs) in a short time (one to two weeks).
REFERENCES
- Takeyama N, Kiyono H, Yuki Y. Plant-based vaccines for animals and humans: recent advances in technology and clinical trials. Ther Adv Vaccines. 2015;3(6):139-154.
[Crossref], [Google Scholar], [PubMed]
- Buyel JF, Fischer R. Predictive models for transient protein expression in tobacco (Nicotiana tabacum L.) can optimize process time, yield, and downstream costs. Biotechnol Bioeng. 2012;109(10):2575-2588.
[Crossref], [Google Scholar], [PubMed]
- Vitale A, Pedrazzini E. Recombinant pharmaceuticals from plants: the plant endomembrane system as bioreactor. Molecular Interventions. 2005 ;5(4):216.
[Crossref], [Google Scholar], [PubMed]
- Hefferon K. Plant‐derived pharmaceuticals for the developing world. Biotech J. 2013;8(10):1193-1202.
[Crossref], [Google Scholar], [PubMed]
- Yusibov V, Rabindran S, Commandeur U, Twyman RM, Fischer R. The potential of plant virus vectors for vaccine production. Drugs in R & D. 2006;7(4):203-217.
[Crossref], [Google Scholar], [PubMed]
- Naseri Z, Khezri G, Davarpanah SJ, Ofoghi H. Virus-based vectors: a new approach for the production of recombinant proteins. J Applied Biotech Rep. 2019;6(1):6-14.
[Crossref], [Google Scholar]
- Bamogo PK, Brugidou C, Sérémé D, Tiendrébéogo F, Djigma FW, Simpore J, et al. Virus-based pharmaceutical production in plants: an opportunity to reduce health problems in Africa. Virol J. 2019;16(1):1-6.
[Crossref], [Google Scholar], [PubMed]
- Patil BL, Fauquet CM. Differential interaction between cassava mosaic geminiviruses and geminivirus satellites. J General Virol. 2010 ;91(7):1871-1882.
[Crossref], [Google Scholar], [PubMed]
- Kumar RV. Plant antiviral immunity against geminiviruses and viral counter-defense for survival. Front Microbiol. 2019;10:1460.
[Crossref], [Google Scholar], [PubMed]
- Veluthambi K, Sunitha S. Targets and mechanisms of geminivirus silencing suppressor protein AC2. Front Microbiol. 2021;12:645419.
[Crossref], [Google Scholar], [PubMed]
- Laufs J, Traut W, Heyraud F, Matzeit V, Rogers SG, Schell J, et al. In vitro cleavage and joining at the viral origin of replication by the replication initiator protein of tomato yellow leaf curl virus. Proc Natl Acad Sci. 1995;92(9):3879-3883.
[Crossref], [Google Scholar], [PubMed]
- Fondong VN. Geminivirus protein structure and function. Mol Plant Pathol. 2013;14(6):635-649.
[Crossref], [Google Scholar], [PubMed]
- Chen Q, He J, Phoolcharoen W, Mason HS. Geminiviral vectors based on bean yellow dwarf virus for production of vaccine antigens and monoclonal antibodies in plants. Hum Vaccin. 2011;7(3):331-338.
[Crossref], [Google Scholar], [PubMed]
- Hefferon KL, Fan Y. Expression of a vaccine protein in a plant cell line using a geminivirus-based replicon system. Vaccine. 2004 Dec 2;23(3):404-410.
[Crossref], [Google Scholar], [PubMed]
- Zhang X, Mason H. Bean yellow dwarf virus replicons for high‐level transgene expression in transgenic plants and cell cultures. Biotechnol Bioeng.2006;93 (2): 271-279.
[Crossref][Google Scholar] [Pubmed]
- Hefferon K L. DNA virus vectors for vaccine production in plants: spotlight on geminiviruses Vaccines 2014; 2 (3): 642-653.
[Crossref][Google Scholar] [Pubmed]
- Atkinson RG, Gleave LR, Janssen BJ, Morris BA. Post‐transcriptional silencing of chalcone synthase in petunia using a geminivirus‐based episomal vector The Plant Journal.1998; 15 (5): 593-604.
[Crossref][Google Scholar] [Pubmed]
- Huang Z, Chen Q, Hjelm B, Arntzen C, Mason H. A DNA replicon system for rapid high‐level production of virus‐like particles in plant. Biotechnol Bioeng.2009; 103 (4): 706-714.
[Crossref][Google Scholar] [Pubmed]
- Huang Z, Phoolcharoen W, Lai H, Piensook K, Cardineau G, Zeitlin L, et.al. High‐level rapid production of full‐size monoclonal antibodies in plants by a single‐vector DNA replicon system. Biotechnol Bioeng.2010; 106 (1): 9-17.
[Crossref][Google Scholar] [Pubmed]
- Regnard GL, Halley‐Stott RP, Tanzer FL, Hitzeroth II, Rybicki EP. High level protein expression in plants through the use of a novel autonomously replicating geminivirus shuttle vector. Plant Biotechnol J.2010; 8 (1): 38-46.
[Crossref][Google Scholar] [Pubmed]
- Diamos AG, Hunter JG, Pardhe MD, Rosenthal SH, Sun H, Foster BC, et.al. High level production of monoclonal antibodies using an optimized plant expression system. Front Bioeng Biotechnol.2020; 7:472.
[Crossref][Google Scholar] [Pubmed]
- Dugdale B, Mortimer CL, Kato M, James TA, Harding RM, Dale DL, et.al. In plant activation: an inducible, hyperexpression platform for recombinant protein production in plants. Plant Cell. 2013; 25 (7): 2429-2443.
[Crossref][Google Scholar] [Pubmed]
- Chen Q. Expression and manufacture of pharmaceutical proteins in genetically engineered horticultural plants. In Transgenic Horticultural Crops: Challenges and Opportunities, by Taylor & Francis Ralph Scorza. 2011:83-124.
- Platis D, Labrou NE. Affinity chromatography for the purification of therapeutic proteins from transgenic maize using immobilized histamine. J Sep Sci. 2008; 31(4): 636-645.
[Crossref][Google Scholar] [Pubmed]
- Phoolcharoen W, Bhoo SH, Lai H, Ma J, Arntzen CJ, Chen Q, et.al. Expression of an immunogenic Ebola immune complex in Nicotiana benthamiana. Plant Biotechnol J.2011; 9 (7): 807-816.
[Crossref][Google Scholar] [Pubmed]
- Buck CB, Pastrana DV, Lowy DR, Schiller JT. Humana press, generation of HPV pseudovirions using transfection and their use in neutralization assays. Methods Mol Med.2005; 119: 445-462.
[Crossref][Google Scholar] [Pubmed]
- Lamprecht RL, Kennedy P, Huddy SM, Bethke S, Hendrikse M, Hitzeroth II, et.al. Production of human papillomavirus pseudovirions in plants and their use in pseudovirion-based neutralisation assays in mammalian cells. Sci Rep. 2016; 6 (1): 1-10.
- Shanmugaraj B, Rattanapisit K, Manopwisedjaroen S, Thitithanyanont A. Monoclonal antibodies B38 and H4 produced in Nicotiana benthamiana neutralize SARS-CoV-2 in vitro. Front Plant Sci.2020; 11: 1-11.
[Crossref][Google Scholar] [Pubmed]
- Moore MJ, Proudfoot NJ. Pre-mRNA processing reaches back totranscription and ahead to translation. Cell.2009; 136 (4): 688-700.
- Diamos AG, Rosenthal SH, Mason HS. 5 and 3 untranslated regions strongly enhance performance of geminiviral replicons in Nicotiana benthamiana leaves. Front Plant Sci. 2016;7:200.
[Crossref], [Google Scholar], [Pubmed]
- Yamamoto T, Hoshikawa K, Ezura K, Okazawa R, Fujita S, Takaoka M, et al. Improvement of the transient expression system for production of recombinant proteins in plants. Sci Rep. 2018;8(1):1-10.
[Crossref], [Google Scholar], [Pubmed]
- Diamos AG, Mason HS. Chimeric 3’flanking regions strongly enhance gene expression in plants. Plant biotechnology journal. 2018;16(12):1971-1982.
[Crossref], [Google Scholar], [Pubmed]
- Garrido-Ramirez ER, Sudarshana MR, Lucas WJ, Gilbertson RL. Bean dwarf mosaic virus BV1 protein is a determinant of the hypersensitive response and avirulence in Phaseolus vulgaris. Mol Plant Microbe Interact .2000;13(11): 1184-1194.
[Crossref], [Google Scholar], [Pubmed]
- Van Wezel R, Dong X, Blake P, Stanley J, Hong Y. Differential roles of geminivirus Rep and AC4 (C4) in the induction of necrosis in Nicotiana benthamiana. Mol Plant Pathol. 2002;3(6):461-471.
[Crossref], [Google Scholar], [Pubmed]
- Diamos AG, Mason HS. Modifying the replication of geminiviral vectors reduces cell death and enhances expression of biopharmaceutical proteins in Nicotiana benthamiana leaves. Front Plant Sci. 2019;9:1974. [Crossref],
[Google Scholar], [Pubmed]
- Kim KI, Sunter G, Bisaro DM, Chung IS. Improved expression of recombinant GFP using a replicating vector based on Beet curly top virus in leaf-disks and infiltrated Nicotiana benthamiana leaves. Plant Mol Biol. 2007;64(1):103-112.
[Crossref], [Google Scholar], [Pubmed]
- Chung HY, Lee HH, Kim KI, Chung HY, Hwang-Bo J, Park JH, et al. Expression of a recombinant chimeric protein of hepatitis A virus VP1-Fc using a replicating vector based on Beet curly top virus in tobacco leaves and its immunogenicity in mice. Plant Cell Rep. 2011;30(8):1513-1521.
[Crossref], [Google Scholar], [Pubmed]
- Chung HY, Park JH, Lee HH, Kim KI, Chung HY, Sunter G, et al. Expression of a functional recombinant chimeric protein of human hepatitis A virus VP1 and an Fc antibody fragment in transgenic tomato plants. Plant Biotechnol Rep. 2014;8(3):243-249.
- Kim KI, Chung HY, Yoo KH, Park JH, Lee HH, Soek YJ, et al. Expression of a recombinant chimeric protein of human colorectal cancer antigen GA733-2 and Fc fragment of antibody using a replicating vector based on Beet curly top virus in infiltrated Nicotiana benthamiana leaves. Plant Biotechnol Rep. 2012;6(3):233-242.
- Kharazmi S, Ataie Kachoie E, Behjatnia SA. Cotton leaf curl Multan betasatellite DNA as a tool to deliver and express the human B-cell lymphoma 2 (Bcl-2) gene in plants. Mol Biotechnol. 2016;58(5):362-372.
- Ataie Kachoie E, Behjatnia SA, Kharazmi S. Expression of Human Immunodeficiency Virus type 1 (HIV-1) coat protein genes in plants using cotton leaf curl Multan betasatellite-based vector. PloS one. 2018;13(1):e0190403.
[Crossref], [Google Scholar], [Pubmed]
- Girard MP, Osmanov SK, Kieny MP. A review of vaccine research and development: the human immunodeficiency virus (HIV). Vaccine. 2006;24(19):4062-4081.
[Crossref], [Google Scholar], [Pubmed]
- Dugdale B, Mortimer CL, Kato M, James TA, Harding RM, Dale JL. Design and construction of an in-plant activation cassette for transgene expression and recombinant protein production in plants. Nat Protoc. 2014;9(5):1010-1027.
[Crossref], [Google Scholar], [Pubmed]
- Matzeit V, Schaefer S, Kammann M, Schalk HJ, Schell J, Gronenborn B. Wheat dwarf virus vectors replicate and express foreign genes in cells of monocotyledonous plants. Plant Cell. 1991;3(3):247-258.
[Crossref], [Google Scholar], [Pubmed]
- Palmer KE, Thomson JA, Rybicki EP. Generation of maize cell lines containing autonomously replicating maize streak virus-based gene vectors. Arch Virol. 1999;144(7):1345-1360.
[Crossref], [Google Scholar], [Pubmed]
- Palmer KE, Rybicki EP. Investigation of the potential of maize streak virus to act as an infectious gene vector in maize plants. Arch Virol. 2001;146(6):1089-1104. [Crossref],
[Google Scholar], [Pubmed]
- Ward A, Etessami P, Stanley J. Expression of a bacterial gene in plants mediated by infectious geminivirus DNA. EMBO J. 1988;7(6):1583-1587.
[Crossref], [Google Scholar], [Pubmed]
Results
In this system, Nicotiana benthamiana leaves were agro infiltrated with a BeYDV replicon vector encoding the Hepatitis B core antigen (HBc) or NVCP along with the Rep/RepA cassette and a p19 vector which is a silencing suppressor protein of Tomato bushy stunt virus. As a result, it got HBc and NVCP at 0.8 and 0.34 mg/g Leaf Fresh Weight (LFW), respectively (Figure 2). These values were reported to be 4-fold more than leaf infiltration with expression constructs without P19 [18]. The three-vector BeYDV replicon system also allows synchronous efficient replication of two distinct genes encoding fluorescence proteins, GFP and DsRed. These proteins were co-expressed as non-competing in up to 80 percent of similar plant cells [19].

Figure 2: Schematic representation of the geminivirus expression constructs. (a) An example of a three-vector replicon system is illustrated in which the Rep gene is expressed in trans. (b) The autonomously replicating vector is shown to contain Rep gene in cis in this construct; (c) An expression viral vector containing optimized genetic elements simultaneously expresses two different proteins; (d) The INPACT cassette expresses the splitting gene only in the presence of the Rep/RepA proteins under the control of the AlcA: AlcR gene switch. GOI: Gene Of Interest; 35S; 35S CaMV promoter, T; terminator; ORFs C1 and C2, encoding Rep and RepA; MAR, the tobacco Rb7 matrix attachment region; PsaK2T 5', the truncated 5' UTR from the N. benthamiana psaK gene; NbACT 3', the 3' UTR from the N. benthamiana ACT3 gene; AlcR, alcohol receptor gene; AlcAP, AlcA promoter. LIR: Long Intergenic Region; SIR: Short Intergenic Region; LB and RB, the Left and Right Borders of the T-DNA region. a, b, c, and d constructs sourced from [18,20-22].
Although the BYDV rep gene provides well transgene replication in trans, efforts to produce a simpler, high-performance system led to the development of a new platform in which vector replication elements were located in cis (Figure 2). The autonomously replicating vector, pRIC, was first constructed from a mild strain of BeYDV in which CP and MP were replaced by the subunit vaccine antigen such as human immunodeficiency virus subtype C p24 or Human Papillomavirus Subtype 16 (HPV-16) major CP L1. The expression of L1 and p24 proteins from pRIC was 50% and three to seven times higher than that from a non-replicative vector, respectively. It was shown that enhanced proteins expression was associated with replication of pRIC in plant cells and that vector replication in Agrobacterium cells or bacterial replication itself was not observed in infiltrated leaf tissue [20].
High-efficiency production of hetero-oligomeric proteins using viral vectors made from the same virus is difficult because the expression and assembly of three or more separate subunit proteins in one cell is ever associated with competing replicons. In addition, it is hard work to find noncompeting viral vectors for the production of multi-segment proteins like Monoclonal Antibodies (mAbs). This problem was resolved using BeYDV system due to the noncompeting nature of its replicons and was able to achieve high performance of both the heavy and light chain subunits of an IgG mAb 6D8 with complete assembly against Ebola Virus Glycoprotein (EBV GP1) (~0.5 mg/g LFW) [20-22]. The host plant is commonly utilized for agro infiltration in N. benthamiana due to its high biomass yield and susceptibility to virus infection [23]. However, nicotiana leaves possess high levels of alkaloids and phenolics, which make the protein purification process difficult [24]. Lai et al. 2012 demonstrated the strong performance of BeYDV vector (pRIC vector) in producing two therapeutic mAbs against EBV and West Nile Virus (WNV) as well as a VLP derived from the NVCP in lettuce (~0.23-0.27 mg/g FLW), which was similar to those of their yield in Nicotiana. Lettuce grows rapidly on a large scale while producing a low level of secondary metabolites, thus making protein purification easier and reducing downstream costs [25]. In another study, the BeYDV vector was exploited to produce Human Papillomavirus (HPV) antibodies using Pseudovirion (PsV)-based Neutralization Assay (PBNA). For this purpose, autoreplicating BeYDV-derived pRIC vector plasmid was modified by inserting a mammalian expression cassette containing a reporter gene called the Secreted Embryonic Alkaline Phosphatase (SEAP) that can be effectively encapsidated into the HPV virion [26]. The PsV were generated by agro infiltration of plants with plasmids expressing SEAP, and capsid proteins (L1 and L2) which showed successful neutralization through known antibodies [27].
Discussion
Recently, two other functional mAbs (B38 and H4) have been produced in plant expression systems that efficiently neutralized Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), the causative agent of the ongoing pandemic coronavirus disease 2019 (COVID-19). These antibodies accumulated in N.benthamiana leaves at levels of 4 and 34 μg/g, respectively [28]. Another way to increase the level of gene expression in the BeYDV system is to use appropriate Untranslated Regions (UTRs) that play an important role in optimizing protein production via RNA stability, efficient transcription processing, and inhibiting gene silencing [29]. Incorporating a truncated of the Arabidopsis thaliana and N. benthamiana photosystem I K (PsaK) 5´UTRs, tobacco Extension terminator (Ext), and the tobacco Rb7 Matrix Attachment Region (MAR) in the 3´ position increase NVCP production more than fourfold the level previously reported in the original vector (1.8 mg/g LFW vs. 0.34 mg/g LFW). Furthermore, the presence of 3´MAR in this system reduced cell death and generated rituximab mAbs at up to 1 mg/g LFW, a 3.4-fold improvement over the vector without MAR. Although the exact function of MAR in the transient expression system is unclear, a hypothesis has been proposed that it may act as a second gene terminator [30]. Recently, Yamamoto et al. demonstrated that introducing a dual terminator consisting of Heat Shock Protein (HSP) and tobacco Ext terminators into the BeYDV replication system increased Green Fluorescent Protein (GFP) expression level in plants (such as nicotiana, lettuce, eggplants, tomato, etc.,), possibly as a result of implicating in the reduction of transgene silencing by RNA-dependent RNA polymerase 6 (RDR6) [31]. The gene terminator and surrounding regions are important segments of viral expression systems, and their improvement helps increase protein production in plants. A study has investigated the potential of terminators from diverse sources in protein expression. Among the tested terminators, the intron less tobacco Extension Terminator (EU) provided a remarkable 13.6-fold increase compared with the Nopaline Synthase (NOS) terminator from A. tumefactions, while the A. thaliana 18.2 kDa Heat-Shock Protein terminator (AtHSP), the potato pin II 3´UTR, and the rubisco small unit (rbcS) 3´UTR from pea increased protein expression by 2.5, 8.5, and 5.4-fold higher, respectively, that of NOS terminator alone. It was identified novel candidates, the 3´ flanking regions from an N.benthamina homolog of the A.thaliana 17.6 kDa class II HSP (At5g12020) and a downstream 617-nt region from an N.benthamina homolog of the A.thaliana actin 7 (Atg09810), referred to as NbHSP and NbACT3, respectively, exhibited greater than 6.3-fold, and 3.9-fold increase. The combination of tandem terminators and MAR dramatically enhanced GFP production, so that constructs consisting of the two best combined dual terminators with Rb7 MAR as the 35S-NbACT3-Rb7 and EU- 35S-Rb7, were developed in a replicating BeYDV vector, displaying >150-fold increase in expression compared to the original NOS vector alone [32]. Further research optimized genetic elements in plant expression BeYDV vectors to simultaneously co-express four fluorescent proteins, so multiple cassettes were create with identical UTRs containing a PsaK 5´UTR, the EU linked to the NbACT3 3´UTR, and Rb7 MAR, which were able to represent similar production levels of each individual proteins without any loss of total yield (Figure 2c). In addition, expression cassettes were generated with suboptimal genetic elements but varied [TMV 5´ with RbcS 3´ and Tobacco Etch Virus (TEV) 5´ with vegetative storage protein B (VspB) 3´] that would be permitted fine-tuning of the expression level of each protein subunits and, was a novel idea for expression of heteromultimeric proteins requiring different proportions of each subunit. These optimized vectors produced three mAbs, 1.47 mg/g LFW 2A10G6 (the variable regions of murine antibody), 1.42 mg/g LFW herpes simplex virus (HSV8), and 1.21 mg/g LFW 6D8 [21]. The Hypersensitive Response (HR) is one factor reducing the yield of recombinant proteins in BeYDV system elicited by viral (in response to virus infection) proteins [33,34]. The overexpression of Rep/RepA, viral replication, and Agrobacterium concentrations contributed to HR. it was reported that by introducing a mutation to the 51 UTR of Rep/RepA, a controlled decrease in their expression was obtained, resulting in reduced leaf cell death and increased mAbs yield. Infiltration of replicating-vector with lower bacteria concentration (OD600 of attained optimum expression0.2) attained optimum expression of proteins, while the higher concentration of Agrobacterium was required (OD600 of ~1.2) using the non-replicating vector [35].
Beet Curly Top Virus (BCTV)
Beet Curly Top Virus (BCTV) is another geminivirus that has been used for the plant virus-based transient expression vector system. This vector is compatible with the application of silencing suppressor p19 and heterologous promoters derived from diverse sources such as the Maize Ubiquitin (MUBI), Cassava Vein Mosaic Virus (CsVMV), Arabidopsis Ubiquitin 3 (AtUBI), and Actin 8 (AtACT). Among the promoters tested, the construct containing the CsVMV promoters showed the highest activity compared to the others, and also supported GFP mRNA accumulation in N. benthamiana at the level 320% higher than the CaMV 35S promoter construct. It was also observed that p19 in BCTV replicon enhances GFP level and was suggested to improve the expression system similar to that of previously reported from a BeYDV vector [36,18]. The recombinant chimeric protein of hepatitis A virus capsid protein VP1 fused to the immunoglobulin Fc fragment (HPV VP1-Fc) has successfully been expressed in transgenic plants (approximately 0.6% total soluble protein from tobacco leaf material and 0.57μg/g from transgenic tomato leaf material) using a BCTVreplicating vector system. This recombinant vaccine antigen driven from the transgenic plants induced a robust specific IgG antibody response in mice after intraperitoneal immunization and also lead to an increase in the secretion of both IFN-y and IL-4 upon immune responses [37,38]. Similar results were obtained for the expression and immunogenicity of recombinant human colorectal cancer antigen GA733-2 fused to the Fc fragment of antibody in Agrobacterium-infiltrated N.benthamiana leaves by using a BCTVbased transient expression vector system. The expression level of GA733-2-Fc was estimated to be about 0.96% of total soluble protein from tobacco leaf material [39].
Cotton Leaf Curl Multan Betasatellite (CLCuMB)
The betasatellites (DNA-βs) have been often isolated from plants infected with the monopartite geminiviruses belonging to genus begomovirus including Cotton Leaf Curl Multan Virus (CLCuMV), and are capable to replicate in plants in the presence of their helper viruses. The DNA-β associated with CLCuMV (CLCuMB) is about 1350 nt in size, comprising a single coding region termed ORF βC1 that is responsible for the induction of disease symptoms in some host plants, an adenine-rich region, and a satellite conserved region. Kharazmi et al. described a new strategy for the expression of human B-cell lymphoma 2 (Bcl-2) in N. benthamiana leaf strips using the CLCuMB. In this method, the recombinant CLCuMB construct was developed by the insertion of Bcl-2 gene (720 nt in size) into a mutant of CLCuMB in which the βC1 (354 nt in size) was deleted. This recombinant viral vector has its promoter and successfully was replicated in tobacco and tomato in the presence of diverse helper geminiviruses including Beet Curly Top Virus (BCTV-Svr), Tomato Yellow Leaf Curl Virus (TYLCV-(Ab)), and Tomato Leaf Curl Virus-Australia (TLCV-Au), as well as was achieved acceptable levels of Bcl-2 gene transcription in these plants. However, it has been reported that some inoculated plants have lost their sensitivity to the viral expression vector, speculating that the size of the introduced sequence may be effective in this stability. It has been suggested that the removal of another region of CLCuMB such as the A-rich region, or the use of only the sequence required for promoter activity, besides the βC1 ORF, may lead to its tolerance to the insertion of the larger foreign gene [40]. Further research showed that a replicating gene expression vector based on CLCuMB expressed the Human Immunodeficiency Virus type 1 (HIV-1) p24 protein at a level of 79-814 ng/LFW in tobacco in the presence of TYLCV-(Ab) infectious construct whereas the recombinant HIV-1 gag protein was not because of its size. The p24 is a 696 bp capsid protein derived from the gag polyprotein with 1051 bp in size [41]. Because both proteins are highly conserved in the HIV-1 genome, they are usually candidates for vaccine production [42].
Tobacco yellow dwarf virus
A novel expression system, In-Plant Activation (INPACT), has been developed based on the rolling circle replication machinery of another mastrevirus, Tobacco Yellow Dwarf Virus (TYDV) that provided a high level of recombinant protein (up to 10% total soluble protein) in plants. The INPACT cassette is uniquely designed so that the gene of interest is split into two exons organizing in a way that exon 1 is placed downstream of exon 2. Both exons are flanked by the TYDV LIRs incorporating within an intron (Figure 2). The INPACT cassette expressed recombinant protein only in the presence of an ethanol-inducible activation cassette encoding the TYDV Rep and RepA replication and processed to remove the intron. This platform is adaptable to a wide range of plant species and also quickly provides low to medium volumes of recombinant protein expression at high levels [22,43].
Other geminivirus vectors
Cassava Latent Virus (CLV), Wheat Dwarf Virus (WDV), and Maize Streak Virus (MSV) are other geminiviruses that are introduced as expression vectors for the investigation of viral gene functions or monitoring viral replication. About these vectors, early expression studies involved the replacement of the virus gene encoding the Capsid Protein (CP) with a reporter gene such as BAR, CAT, GFP, and GUS [44-47]. For example, a deletion of 727 nucleotides within the CLV CP gene has been substituted by the coding region of the CAT gene show that the CP plays no necessary role in virus spread entirely the plant [47].
Conclusion
Using plant viruses as expression vectors lead to the production of vaccines and therapeutic complex proteins in a rapid, inexpensive, high- scalability, and safe manner. Geminiviruses offer several advantages, including the ability to express foreign proteins in a border range of plants, their genome is easy to manipulate, so it takes a short time to develop a product that is very useful in fighting the global pandemic. Future research will focus on optimizing and improving the design of vectors and their function to produce high efficiency with biosafety.
Citation: lotfipour M (2022) Recent Advances for Vaccine Production in Plants Using Geminiviruses. J Vaccines Vaccin. 13:476.
Copyright: © 2022 lotfipour M. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.