
Indexed In
- Academic Journals Database
- Genamics JournalSeek
- Academic Keys
- JournalTOCs
- China National Knowledge Infrastructure (CNKI)
- Scimago
- Access to Global Online Research in Agriculture (AGORA)
- Electronic Journals Library
- RefSeek
- Directory of Research Journal Indexing (DRJI)
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- SWB online catalog
- Virtual Library of Biology (vifabio)
- Publons
- MIAR
- University Grants Commission
- Geneva Foundation for Medical Education and Research
- Euro Pub
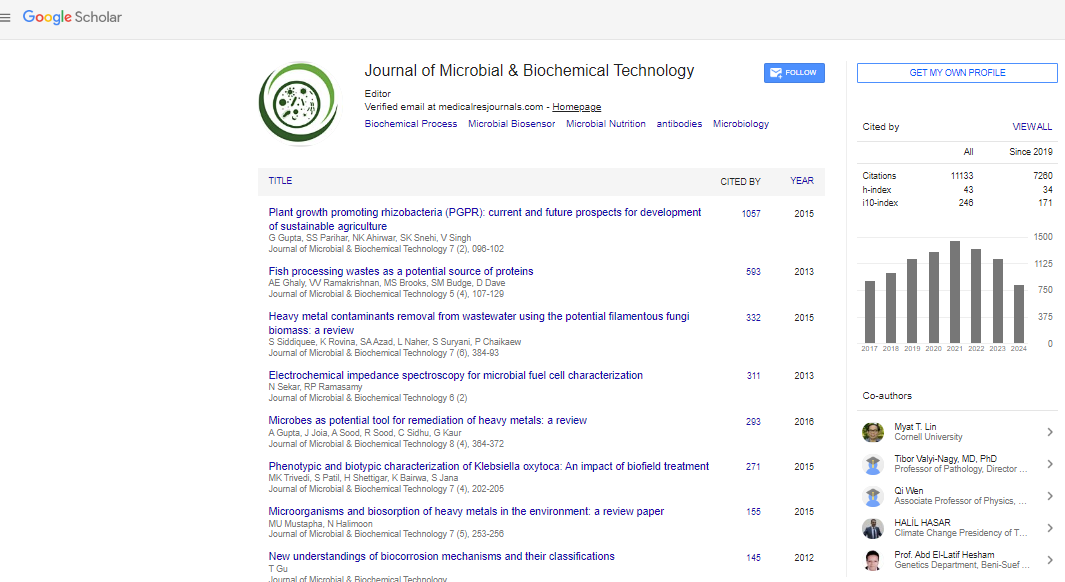
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Research - (2021) Volume 13, Issue 6
Microbial Technology Agriculture
Received: 01-Jun-2021 Published: 24-Jun-2021, DOI: 10.35248/1948-5948.21.13.476
Abstract
Food production has been increased as a result of the fast adoption of biotechnological techniques that allow for the quick discovery of new chemicals and microbes, as well as the genetic enhancement of established species. Microorganisms have never been more prevalent in fields such as agriculture and medicine in history, save as wellknown villains. Currently, however, different agricultural crops require helpful microbes such as plant growth promoters and controllers for phytopathogens, and many species are utilised as bio-factories for essential pharmaceutical compounds.
Introduction
Compounds having pesticidal action, primarily herbicidal, insecticidal, and nematicidal, have recently piqued researchers' interest in microorganisms. To manage Morrenia odorata, the first commercially licensed mycoherbicide was a suspension of Phytophthora palmivora and numerous more plant parasite and phytotoxin-producing microbial species have subsequently been found [1]. Colletotrichum gloeosporioides can cause anthracnose symptoms in Aeschynomene virginica, a rice and soybean weed, allowing it to be controlled. By totally suppressing blooming and decreasing tuber development, Puccinia canaliculata can control yellow nutsedge (Cyperus esculentus L) [2].
Bio-herbicides, on the other hand, have not been widely used in agronomic and horticultural crops for weed control since they require a variety of parameters, such as optimum humidity, that reduce their efficacy when compared to conventional herbicides. Biotechnological advancements will most likely reverse this trend and increase the performance of bio-herbicides in the future.
Cry and Cyt, two endotoxin proteins, are now used as insecticides. The soil bacteria B. thuringiensis (Bt) produces these endotoxins, which have an entomopathogenic effect on pests found in cabbage, potatoes, and cereals [3]. Several transgenic plants producing Bt protein, eg. Maize, tobacco, and tomato, have been grown across the world because to their efficacy in preventing caterpillars, particularly Lepidoptera, from spreading [4]. Baculovirus may infect caterpillars and eggs of pests like Spodoptera frugiperda, decreasing agricultural losses caused by this insect, notably in maize. Furthermore, the development made in the virus's genetic modification has enhanced its efficiency as an insecticide [5]. Several fungi harmful to insects, such as Beauveria, Metarhizium, and Paecilomyces, are also being utilised as control agents. These are most commonly employed to combat leaf caterpillars in greenhouses or other locations with high humidity [6].
Much progress has been made in the research and marketing of bionematicides in recent years [7]. The metabolites known as avermectins produced by the bacteria Streptomyces avermitilis are an example of this. These pesticides are model pesticides because they are non-toxic to mammals and active against nematodes at extremely low concentrations. Therefore, B. firmus culture filtrates are used against adult nematodes and larvae; majorly Radopholus similis, Meloidogyne incognita, and Ditylenchus dipsaci. This filterate paralyse these nematodes and kill them, which suggest that toxic metabolite synthesis is involved in the control of these pests [8]. When Myrothecium verrucaria is cultivated in bioreactors, toxic metabolites are generated, and when these metabolites come into contact with adult nematodes, the metabolites in suspension kill the adults while also preventing egg development and hatching [9]. Pasteuria sp. endospores, on the other hand, employ parasitism as a means of control. When these endospores come into touch with nematodes, they germinate, become parasitic, and have a significant impact on host reproduction [10]. The fungus belonging to the genus Trichoderma are the most extensively distributed microorganisms that operate in biological pest management. These fungi are saprophytes, mycoparasite decomposers, and plant symbionts that are often found in soil environments and have a worldwide geographical range [11]. This diversity of lifestyles within the genus explains why Trichoderma is the source of numerous strains employed in biological control. Trichoderma spp. parasitise and suppress a wide range of phytopathogenic fungi and exhibit a nematicidal effect on the Meloidogyne [12]. This functional feature of Trichoderma and other species answer back to the growing need for methods that reduce pesticide side effects such as insect population resistance, soil and water quality decrease, and the formation of residues with negative effects on non-target organisms. Sustainable agriculture, on the other hand, includes not only the management of phytopathogens but also the utilisation of functional microbial features associated to plant growth promotion. Mycorrhizal fungi and rhizobacteria, for example, produce activities that can increase plant fitness by enabling nutrient absorption by the plant. Mycorrhizal fungi and roots complement each other in plant foraging inside nutrient patches [13] and help the plant acquire phosphorus by expressing genes that code for inorganic phosphorus transporters [14]. Plant growth-promoting rhizobacterias (PGPRs), too, stimulate plant development through direct and indirect processes. Direct methods include nitrogen synthesis by strains belonging to the genera Rhizobium, Allorhizobium, Azorhizobium, Bradyrhizobium, Mesorhizobium, and Sinorhizobium, as well as root growth stimulation via auxin, cytokinin, and gibberellin production. Antibiosis, development of systemic resistance, and competition for resources and habitats are examples of indirect processes that reduce disease vulnerability [15].
Endophytic bacteria, on the other hand, invade plant tissues without causing disease symptoms, creating a stable long-term relationship with the host plant. Endophytes produce bioactive compounds during the contact, which may improve the plant's fitness. Nitrogen fixation, phytohormone production, biocontrol of phytopathogens through the manufacture of antibiotics or siderophores, competition for nutrients, and the creation of systemic disease resistance may all contribute to endophytic growth promotion [16]. However, the bioprospection and identification of these microbes associated with a wide range of plant species is not only important to obtain the strains of agricultural importance, but also to identify such strains those can produce bio-products having antibiotic ability [17], as well as potential for achieving biotechnologically important chemicals [17].
REFERENCES
- McRae CF. Classical and inundative approaches to biological weed control compared. Plant Prot. Quart. 1988;3:124–127.
- Duke SO, Scheffler BE, Boyette CD, Dayan FE. Biotechnology in weed control. Kirk-Othmer Encyclopedia of Chemical Technology. New York, NY: John Wiley & Sons, Inc. 2015.
- Sarwar M. Biopesticides: an effective and environmental friendly insect-pests inhibitor line of action. Int. J. Eng. Adv. Res. Technol. 2015;1: 10–15.
- Khan MA, Paul B, Ahmad W, Paul S, Aggarwal C, Khan Z, et al. Potential of Bacillus thuringiensis in the management of pernicious lepidopteran pests, in Plant, Soil and Microbes, eds K. R. Hakeem and M. S. Akhtar (New York City, NY: Springer International Publishing), 2016; 277–301.
- Popham HJR, Nusawardani T, Bonning BC. Introduction to the use of baculoviruses as biological insecticides, in Baculovirus and Insect Cell Expression Protocols, ed. D.W. Murhammer (New York, NY: Springer), 2016;383–392.
- Sarwar M. Biopesticides: an effective and environmental friendly insect-pests inhibitor line of action. Int. J. Eng. Adv. Res. Technol. 2015;1: 10–15.
- Wilson MJ, Jackson TA. Progress in the commercialisation of bionematicides. BioControl. 2013;58: 715–722.
- Mendoza AR, Kiewnick S, Sikora RA. In vitro activity of Bacillus firmus against the burrowing nematode Radopholus similis the root-knot nematode Meloidogyne incognita and the stem nematode Ditylenchus dipsaci. Biocontrol. Sci. Technol. 2008;18: 377–389.
- Twomey U, Warrior P, Kerry BR, Perry RN. Effects of the biological nematicide, DiTera, on hatching of Globodera rostochiensis and G. pallida. Nematology. 2000;2: 355–362.
- Davies KG, Rowe J, Manzanilla-López R, Opperman CH. Re-evaluation of the life-cycle of the nematode-parasitic bacterium Pasteuria penetrans in root-knot nematodes. Meloidogyne spp. Nematologica. 2011;13: 825–835.
- Druzhinina IS, Seidl-Seiboth V, Herrera-Estrella A, Horwitz BA, Kenerley CM, Monet E, et al. Trichoderma: the genomics of opportunistic success. Nat. Rev. Microbiol. 2011; 9: 749–759.
- Sokhandani Z, Moosavi MR, Basirnia T. Optimum concentrations of Trichoderma longibrachiatum and cadusafos for controlling Meloidogyne javanica on Zucchini plants. J. Nematol. 2016;48: 54–63.
- Cheng L, Chen W, Adams TS, Wei X, Li L, McCormack ML, et al. Mycorrhizal fungi and roots are complementary in foraging within nutrient patches. Ecology. 2016;97: 2815–2823.
- Walder F, Boller T, Wiemken A, Courty PE. Regulation of plants’ phosphate uptake in common mycorrhizal networks: role of intraradical fungal phosphate transporters. Plant Signal. Behav. 2016;11:1131-1372.
- Lugtenberg B, Kamilova F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009;63: 541–556.
- Ahmad S, Abbas SS, Prakash R, Alam A, Husain MA. Applications of endophytic Actinomycetes and their role in protection. Imp. J. Interdiscip. Res. 2016;2: 854–858.
- Ma YM, Liang XA, Zhang HC, Liu R. Cytotoxic and antibiotic cyclic pentapeptide from an endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016;64: 3789–3793.
Citation: Choubey P. (2021) Microbial Technology in Agricultural. Journal of Microbial & Biochemical Technology. doi: 13:.6.35248/1948- 5948.21.13.476.
Copyright: © 2021 Choubey P. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.