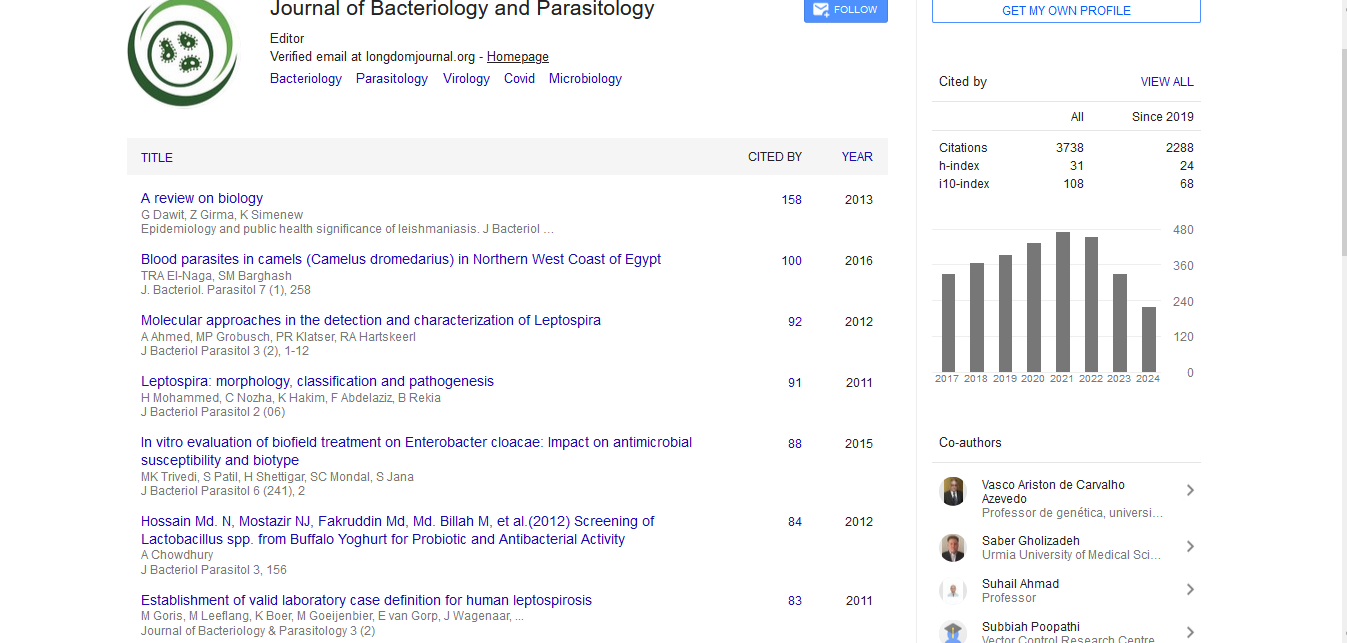

Indexed In
- Open J Gate
- Genamics JournalSeek
- Academic Keys
- JournalTOCs
- ResearchBible
- Ulrich's Periodicals Directory
- Access to Global Online Research in Agriculture (AGORA)
- Electronic Journals Library
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- SWB online catalog
- Virtual Library of Biology (vifabio)
- Publons
- MIAR
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Mini Review - (2020) Volume 11, Issue 4
Free Flavin Participates in Iron and Also Oxygen Metabolism in Bacteria
Tomonori Suzuki1, Shinya Kimata1, Junichi Satoh1, Kouji Takeda2, Daichi Mochizuki1, Ken Kitano3, Akio Watanabe1, Etsuo Yoshimura4, Masataka Uchino1, Shinji Kawasaki1, Akira Abe1 and Youichi Niimura1*2Education course, Tokyo University of Agriculture, Tokyo, Japan
3Nara Institute of Science and Technology, Ikoma, Japan
4The Open University of Japan, Chiba, Japan
Received: 11-Jul-2020 Published: 28-Jul-2020, DOI: 10.35248/2155-9597.20.11.377
Abstract
The reduction of ferric iron to ferrous iron is an essential reaction for the utilization of iron in vivo and ferric reductase participate in this reduction. There are two types of ferric reductase reactions: a reaction using free flavin (non-protein-bound flavin), and a reaction independent of free flavin. However, most of all ferric reductase activities tested to date, including both types of ferric reductases, are enhanced by the addition of free flavin. In E. coli, the iron release from iron storage proteins is markedly stimulated by both types of ferric reductases in the presence of free flavin. Free flavins can directly carry electrons toward molecular oxygen in the aerobic metabolism of an aerotolerant anaerobe, Amphibacillus xylanus. The free flavin and its associated system are able to participate in oxygen and iron metabolism during aerobic bacterial growth. The reaction products of reduced free flavin with oxygen and Fe3+ cause the Fenton reaction, producing the most cytotoxic hydroxyl radical, •OH. Finally, the contribution of free flavin to the process of the Fenton reaction is discussed here.
Keywords
Amphibacillus xylanus; E. coli; Free flavin; Fenton reaction; Ferric reductase
Introduction
Flavin adenine dinucleotide (FAD), flavin mononucleotide (FMN), and riboflavin are the general flavin species present in cells. FAD and FMN bind to flavoproteins or flavoenzymes, and function as their cofactors. These proteins or enzymes function not only in redox reactions but also in electron transport. However, several biological reactions that require non-protein binding flavins (free flavins: free FAD or free FMN) have been reported in bacterial cells (Figure1), [1-19]. In those reports, the free flavins predominantly participated in iron metabolism in bacteria.

Figure 1: Free flavin and protein binding flavin.
Iron is an essential metal in biological processes such as DNA synthesis, intracellular respiration, nitrogen fixation, and photosynthesis, in which it is utilized as a cofactor for ironbinding proteins. It is believed that iron-binding proteins may Journal of Bacteriology & Parasitology incorporate iron in the form of ferrous ion. Iron generally, however, exists in its ferric form under normal aerobic conditions. Thus, the reduction of ferric iron (Fe3+) to ferrous iron (Fe2+) is an essential reaction for the utilization of iron in vivo. Free flavins is involved in the reduction of the iron to form of iron-binding proteins. The free flavin participating in iron metabolism is shown in Section 1.
The iron-binding proteins, cytochromes, act to carry electrons from dehydrogenase systems toward molecular oxygen in a respiratory chain. By contrast, free flavins can directly carry electrons toward molecular oxygen in the aerobic metabolism of an aerotolerant anaerobe, Amphibacillus xylanus (Section 2). In the reaction of reduced free flavin with oxygen and Fe3+, H2O2 and Fe2+ , respectively, are formed as products. These reaction products cause the Fenton reaction producing hydroxyl radical, •OH. We focus on the participation of free flavin in the process of Fenton reaction in Section 3.
Free flavin participates in iron metabolism
There are two types of ferric reductase reactions which are an essential reaction for the utilization of iron in vivo: although a large number of free flavin (non-protein binding flavin) dependent ferric reductases have been reported in many organisms [3,10,12,15,19-27]. Some free flavin independent ferric reductases have also been done [25,28,29].
We purified the enzymes showing free flavin-independent ferric reductase activity from E. coli, ferredoxine-NADP+ reductase (Fpr) and oxygen insensitive nitroreductase (NfnB) [16], and from Synechocystis, DrgA protein [14] and ferredoxine-NADP+ reductase (FNR) [17]. Although these purified enzymes showed free flavin-independent ferric reductase activity for ferric iron compounds bound to synthetic chelators, such as Fe(III)-EDTA, these enzymatic activities for the ferric iron compounds bound to natural chelators such as ferric citrate were much lower than those for synthetic chelators. The ferric reduction including natural chelators by these ferric reductases including both types of ferric reductases was enhanced in most cases by the addition of free flavin.
The reduction of Fe3+ in iron storage proteins is an important metabolic reaction for leading to the release of Fe2+ [30]. Some biological low molecular weight reductants such as thiols, ascorbate, and reduced free flavins are known to be involved in the release of ferrous iron from iron storage proteins [8,9]. Especially, reduced free flavins are effective at releasing Fe2+ from the iron storage proteins and is enzymatically generated from its oxidized form using NAD(P)H as a reducing reagent [8]. In our previous study, it was found that Fpr and NfnB purified from E. coli as free flavin-independent ferric reductases have flavin reductase activity [16]. Thus, these findings suggested the possibility that Fpr and NfnB accelerate the release of Fe2+ from iron storage protein in the presence of free flavin.
Actually, natural iron storage proteins prepared from E. coli, ferritin A (FtnA) and bacterioferritin (Bfr), showed the effective Fe2+ release from them by Fpr and NfnB in the presence of free flavins [31]. Fpr and NfnB showed flavin reductase activity for FAD, FMN and riboflavin, and their Fe2+ release activities were positively associated with the catalytic efficiencies (kcat/Km) for individual flavins. The Fe2+ release activity of E. coli cell-free extracts was affected by flavin reductase activity of the extracts. The Butyl TOYOPEARL column chromatography of the extracts, on the basis of NAD(P)H-dependent flavin reductase activity, resulted in the separation of six active fractions containing Fpr, NfnB, NAD(P)H-quinone oxidoreductase (QOR), flavin reductase (Fre) or alkyl hydroperoxide reductase subunit F (AhpF) as major components. Like Fpr and NfnB, recombinant QOR, Fre, and AhpF showed flavin reductase activity and ferrous iron release activity only in the presence of free flavins, indicating an association of free flavin and flavin reductase activity with Fe2+ releasing activity in E. coli. Thus, both types of free flavin dependent and independent ferric reductases need free flavin for the Fe2+ release activity from natural iron storage proteins in E. coli. Among the four enzymes tested in this study, rFpr showed the highest observed rate constants for Fe2+ release from rFtnA and rBfr in the presence of free flavins. Comparatively, Weeratunga et al. and Yao et al. reported that Pseudomonas aeruginosa Fpr (PaFpr) showed the release of Fe2+from iron storage proteins in the absence of free flavin [32,33]. The rate constant for Fe2+release of Fpr in the presence of free flavin was much higher than that of PaFpr in the absence of free flavin.
Taken together, as free flavins are required for releasing the iron stored in ferritin in E. coli, reduced flavins could serve as an efficient reductant for generating Fe2+ in vitro. Thus, it is important to determine the cellular free flavin content of E. coli and other bacteria to understand how the free flavins are involved in the release of Fe2+ from iron storage proteins. Recently, we established a quantitative analysis method for analyzing the composition and concentration of intracellular free flavins in Amphibacillus xylanus [23]. In the near future, the biological function of intracellular free flavins in the iron release mechanism and metabolism may be elucidated using this new method.
Free flavin also participates in oxygen metabolism
The reduction of oxygen by reduced free flavin is a well-known reaction and is essentially irreversible [34]. Intracellular free flavins have been evaluated in E. coli [35]. However, their existence in the cells of bacteria has not been validated because of the lack of effective methods for distinguishing between flavins in their free form and protein-binding form. We previously separated free flavin from protein-binding flavin in A. xylanus using a gel-filtration column [36-39]. However, this method was inadequate for quantification, because free flavins nonspecifically interacted with the column carriers, leading to variations in the yield of free flavins. A quantitative method for determining the concentrations of free flavins was established in A. xylanus applying an ultrafiltration membrane (Figure 2) [23].

Figure 2: Preparation steps for quantification of intracellular free flavins in A. xylanus.
Amphibacillus xylanus was isolated as a biomass utilization bacterium that can assimilate xylan under alkaline conditions in the presence or absence of oxygen [36]. This bacterium showed unique phenotypic and chemotaxonomic characteristics; therefore, it was accepted as a new genus (Amphibacillus) and species (xylanus) [37,38]. Amphibacillus xylanus grows well under aerobic and anaerobic conditions, exhibiting the same growth rate and cell yields under both conditions, despite its lack of a respiratory chain (Figure 3) [23,40].

Figure 3: Anaerobic and aerobic metabolic pathways in A. xylanus.
The aerotolerant anaerobe, A. xylanus, can be precisely distinguished from the typical facultative aerobe, E. coli, having a respiratory chain (Table 1). However, the both of A. xylanus and E. coli consumed oxygen at rate of 130 – 140 μmol oxygen/min/mg dry cells at 37°C. The rate for E. coli largely depends on a respiratory chain which is lacking in A. xylanus. This indicates that A. xylanus has an alternative system that consumes a large amount of oxygen. A. xylanus NADH oxidase (Nox) was previously reported to rapidly reduce molecular oxygen content in the presence of exogenously added free flavin [1,39].
| Group (Species) | Growth | Respiratory chain | Protection enzymes for reactive oxygen species | ||
|---|---|---|---|---|---|
| Anaerobic/ Aerobic | Nox- or AhpF- Prx (AhpC) | Catalase | Peroxidase | ||
| Aerotolerant anaerobe (A. xylanus) | +/+* | - | + | - | - |
| Faculative aerobe (E. coli) | +/+** | + | + | + | + |
*:Similar growth rates were obtained in aerobic and anaerobic conditions.
**: The growth rate under aerobic condition was larger than that under aerobic condition.
Table 1: Differentiation of A. xylanus and E. coli.
By using the established new method for free flavins, the intracellular concentrations of free FAD, free FMN, and riboflavin were estimated to be approximately 8, 3, and 1 μM, respectively [23]. In the presence of the predominant free flavin species FAD, two flavoproteins, Nox, which binds FAD, and NAD(P)H oxidoreductase (Npo), which binds FMN, were identified as central free flavin-associated enzymes in the oxygen metabolic pathway. Under 8 μM free FAD, the catalytic efficiency (kcat/Km) of recombinant Nox and Npo for oxygen increased by approximately 5- and 9-fold, respectively [23]. Nox and Npo levels were increased and intracellular FAD formation was stimulated following exposure of A. xylanus to oxygen [23]. This suggests that these two enzymes and free FAD contribute to effective oxygen detoxification and NAD(P)+ regeneration to maintain redox balance during aerobic growth. Furthermore, A. xylanus required iron to grow aerobically.
A. xylanus lacks haem synthesis genes, but conserves certain genes that express iron-associated proteins, two of which are ferredoxin (fer) and 7 Fe ferredoxin (fdxA) genes. These genes are upregulated under 21% oxygen [40]. In this study, we found that A. xylanus required iron during aerobic growth. Correspondingly, the mRNA expression of the iron-containing enzyme gene encoding peptide deformylase (PDF) was also induced. PDF is essential for the maturation of polypeptides in bacteria. However, it becomes inactive upon exposure to molecular oxygen due to binding of ferrous ion [41]. Thus, continuous expression of PDF holo-enzyme may be required during aerobic growth of A. xylanus.
Iron-binding proteins, including PDF, always incorporate iron in the form of ferrous ion [10]. Therefore, ferric reductase activity is important for activation of these enzymes. The CFE from A. xylanus showed NAD(P)H-dependent ferric reductase activities, which were increased by 2–3-fold in the presence of 8 μM free FAD. A free flavin-dependent increase in ferric reductase activity was also observed in CFEs from other bacteria such as E. coli and several organisms, and flavin reductases were purified as contributors to this reaction [10,14-17]. Nox and Npo, which function as major flavin reductases, showed markedly low ferric reductase activities for ferric citrate and four other synthetic iron compounds. However, their activities towards all substrates were increased after the addition of 8 μM free FAD. This prominent increase in ferric iron reductase activity may also be responsible for reduced free FAD (FADH2), as proposed by Coves and Fontecave [10]. Thus, our results suggest that Nox and Npo participate in iron metabolism and oxygen metabolism in the presence of free FAD in A. xylanus.
In this study, we confirmed that free flavins are present in A. xylanus, and FAD is a predominant free flavin species. The physiological concentration of free FAD was sufficient to stimulate molecular oxygen and ferric iron reductase activities of two flavoproteins, Nox and Npo. A. xylanus grows well under aerobic conditions using glucose as an energy source, but the energy metabolic system predicted in our previous works lacked an efficient NAD(P)H reoxidation system [39,40]. NAD(P)H should be oxidized to NAD(P)+ constantly because NAD(P)+ is reused as a substrate in a glycolytic pathway, pyruvate metabolic pathway, and pentose phosphate pathway for the next cycle of metabolism. Based on our results, the free flavin-associated enzyme system plays a role in the effective NAD(P)H reoxidation system by passing reducing equivalents of NAD(P)H to molecular oxygen or ferric iron via the reduced form of flavin. Thus, in conclusion, the free flavin-associated system can also participate in oxygen metabolism and supplying Fe2+ during aerobic growth in an aerotolerant anaerobe A. xylanus as shown in Figure 3. We previously isolated an aerotolerant anaerobe, Lactobacillus plantarum P1-2, that can efficiently reduce fatty acid hydroperoxides to their corresponding hydroxyl derivatives [42], and found also a free flavin and its associated enzyme for oxygen metabolism in the isolate (Niimura, unpublished data), suggesting that a free flavin-associated system could participate in oxygen metabolism in an aerotolerant anaerobe lacking a respiratory chain.
Free flavin participates in fenton reaction?
We purified 5 enzymes which are capable of functioning as ferric reductase and of driving the Fenton reaction [14-17]. Fpr and NfnB were purified from E. coli; DrgA protein and FNR were from Synechocystis; aldehyde reductase was from Chlorella [14-17]. The Fenton activities of these enzymes in the presence of synthetic chelate iron compounds were higher than those in the presence of natural chelate iron compounds. The ferric reduction by these enzymes was enhanced in most cases by the addition of free flavin. In the enhanced reaction, reduced free flavin made Fe3+ into Fe2+ and O2 into H2O2. The simultaneous production of H2O2 and Fe2+ in cells may promote the production of hydroxyl radical via the Fenton reaction that causes damage to proteins, lipids and DNA, and results in cell death. Accordingly, both H2O2 and ferrous iron must be properly and safely metabolized in vivo.
The oxygen metabolic enzyme in A. xylanus, NADH oxidase, which catalyzes the reduction of oxygen to hydrogen peroxide with NADH, can also reduce hydrogen peroxide to water in the presence of AhpC (Prx) protein, a member of the peroxiredoxin (Prx) family [43-45]. NADH oxidase thus belongs to a family of peroxiredoxin (Prx) oxidoreductases, which also includes alkyl hydroperoxide reductase F (AhpF) [44,45]. The amino acid sequence of the NADH oxidase exhibits 51.2% shared identity with AhpF from Salmonella typhimurium [43,46]. The Km values of both enzymes for the substrates hydrogen peroxide and cumene hydroperoxide are too low to determine using the employed analytical methods [44]. The turnover numbers of the peroxide reductions catalyzed by both enzymes are extremely high compared with those of other known peroxide scavenging enzymes [47-53]. While two distinct proteins take part in the reaction, the NADH oxidase–Prx system can nevertheless reduce hydroperoxides at a similar rate constant for the first step of the enzyme reaction, suggesting that NADH oxidase and Prx interact very closely to reduce hydroperoxides [47,54,55]. The systems are widely distributed in bacterial obligate aerobes, facultative aerobes, aerotolerant anaerobes, and obligate anaerobes [56].
Facultative aerobe E. coli has catalase and AhpF-AhpC (Prx) which contribute to effective hydroperoxides detoxification [57]. E. coli has additional reactive oxygen species protective systems, and also enterobactin as an iron chelating compound. It was found that the flavin-independent Fenton activity by Escherichia coli ferredoxin-NADP+ reductase which may function as a ferric reductase in vivo is lower in the presence of enterobactin than in the presence of other natural chelate iron compounds and synthetic chelate iron compounds [16,58]. Thus, E. coli is protected against the Fenton reaction by the two H2O2- scavenging enzymes and stable Fe2+ chelator. On the other hand, although A. xylanus has a NADH oxidase–Prx system, it lacks catalase and general iron chelators such as siderophores. However, the two bacteria show phenotypic similarity to oxygen and iron, because they grow well in aerobic condition, require iron to grow aerobically and exhibit nearly the same oxygen consumption activity each other. In E. coli, the Fenton reaction has been shown to occur through the reduction of Fe3+ by reduced free flavin produced by flavin reductase in a hyper- reductive environment induced by the respiratory chain blockage and exposure of H2O2 [13]. Thus, to better understand the contribution of free flavin to the process of the Fenton reaction in bacteria, the dynamic states of Fe2+, H2O2 and free flavin in the living bacterial cells should be investigated.
Conclusion
Free flavins accelerate most of all ferric reductase reactions that are essential for the utilization of iron in vivo. Especially, the free flavins are required for releasing the iron stored in ferritin in a facultative aerobe, E. coli, having a respiratory chain. On the other hand, in an aerotolerant anaerobe, A. xylanus lacking a respiratory chain, free flavin participates in the reduction of both of Fe3+ and oxygen . The reaction of reduced flavins with oxygen and Fe3+ produces H2O2 and Fe2+, respectively, which could cause the Fenton reaction producing cytotoxic hydroxyl radical. In most cases, the activities of 5 ferric reductase-like enzymes purified from microorganisms such as E. coli, Synechocystis and Chlorella are capable of driving the Fenton reaction and are enhanced by the addition of free flavin. Accordingly, to maintain the normal state of cells, the reaction products inducing the Fenton reaction, H2O2 and ferrous iron, must be safely metabolized in vivo. The current findings support the participation of free flavins in the bacterial iron and oxygen metabolism. However, further investigations for the kinetics and dynamics of free flavins in various bacteria will be required to confirm it.
Acknowledgement
We thank Prof. Junichi Nakagawa of Tokyo University of Agriculture for helpful discussion. We greatly appreciate late Prof. Vincent Massey at the University of Michigan Medical School for providing valuable advice. We thank Shun Iwakame, Shinya Tatsumi, Enji Morita, Keita Hara, Shun Tomizuka, Tetsuya Nasukawa, Haruto Arima, and Yui Ohno at Tokyo University of Agriculture for helpful technical assistance in experiments of A. xylanus.
We also thank Takanori Fujita, Kazuyuki Gotoh, Hiroyuki Tajima, Hiroaki Kurihara, Rika Nishiyama, Naohito Yoneyama, Eisuke Tanaka, Tetsuya Mori, Aimi Kawabata, Sayuri Yumoto, Shota Nakamoto, Tatsuya Ishii, Kota Sekino at Tokyo University of Agriculture for helpful technical assistance in experiments of Shinechocystis, Chlorella and E. coli. This work was supported by Grant in Aid for Young Scientists (B) Grant Number 24780081 for J. S. and for Scientific Research (C) Grant Number 19K05780 for Y. N. from MEXT.
REFERENCES
- Nishiyama Y, Massay V, Takeda K, Kawasaki S, Sato J, Watanabe T, et al. Hydrogen peroxideforming NADH oxidase belonging to the peroxiredoxin oxidoreductase family: existence and physiological role in bacteria. J Bacteriol. 2001;183(8):2431–2438.
- Fontecave M, Eliasson R, Reichard P. NAD(P)H:flavin oxidoreductase of Escherichia coli. A ferriciron reductase participating in the generation of the free radical of ribonucleotide reductase. J Biol Chem. 1987;262(25):12325-12331.
- Covès J, Nivière V, Eschenbrenner M, Fontecave M. NADPH-sulfite reductase from Escherichia coli. A flavin reductase participating in the generation of the free radical of ribonucleotide reductase. J Biol Chem. 1993;268(25):18604-18609.
- Thibaut D, Ratet N, Bisch D, Faucher D, Debussche L, Blanche F, et al. Purification of the twoenzyme system catalyzing the oxidation of the Dproline residue of pristinamycin IIB during the last step pristinamycin IIA biosynthesis. J Bacteriol. 1995;177(18):5199-5205.
- Blanc V, Lagneaux D, Didier P, Gil P, Lacroix P, Crouzet J, et al. Cloning and analysis of structural genes from Streptomyces pristinaespiralis encoding enzymes involved in conversion of pristinamycin IIB to pristinamycin IIA (PIIA): PIIA synthase and NADH: riboflavin 50-phosphate oxidoreductase. J Bacteriol. 1995;177(18):5206-5214.
- Kendrew SG, Harding SE, Hopwood DA, Marsh NG. Identification of a flavin: NADH oxidoreductase involved in the biosynthesis of actinorhodin. Purification and characterization of the recombinant enzyme. J Biol Chem. 1995;270(29):17339-17343.
- Parry RJ, Li W. An NADPH:FAD oxidoreductase from the valanimycin producer, Streptomyces viridifaciens. J Biol Chem. 1997;272(37):23303-23311.
- Sirivech S, Frieden E, Osaki S. The release of iron from horse spleen ferritin by reduced flavins. Biochem J. 1974;143(2):311-315.
- Melman G, Bou-Abdallah F, Vane E, Maura P, Arosio P, Melman A, et al. Iron release from ferritin by flavin nucleotide. Biochim Biophys Acta. 2013;1830(10):4669-4674.
- Coves J, Fontecave M. Reduction and mobilization of iron by a NAD(P)H:flavin oxidoreductase from Escherichia coli. Eur J Biochem. 1993;211(1):635-641.
- Fontecave M, Covès J, Pierre J. Performance of Radiologists in Differentiating COVID-19 From Viral Pneumonia on Chest CT. Biometals. 1994;7(2):3-8.
- Pierre JL, Fontecave M, Crichton RR. Chemistry for an essential biological process: the reduction of ferric iron. Biometals. 2002;15(1):341-346.
- Woodmansee AN, Imlay JA. Reduced flavins promote oxidative DNA damage in non-respiring Escherichia coli by delivering electrons to intracellular free iron. J Biol Chem. 2002;277(37):34055-34066.
- Takeda K, Iizuka M, Watanabe T, Nakagawa J, Kawasaki S, Niimura Y, et al. Synechocystis DrgA protein functioning as nitroreductase and ferric reductase is capable of catalyzing the fenton reaction. FEBS J. 2007;274(5):1318-1327.
- Sato J, Takeda K, Nishiyama R, Fusayama K, Arai T, Sato T, et al. Chlorella vulgaris aldehyde reductase is capable of functioning as ferric reductase and driving the fenton reaction in the presence of free flavin. Biosci Biotechnol Biochem. 2010;74(4):854-857.
- Takeda K, Sato J, Goto K, Fujita T, Watanabe T, Abo M, et al. Escherichia coli ferredoxin-NADP+ reductase and oxygen-insensitive nitroreductase are capable of functioning as ferric reductase and of driving the Fenton reaction. Biometals. 2010;23(1):727-737.
- Sato J, Takeda K, Nishiyama R, Watanabe T, Abo M, Yoshimura E, et al. Synechocystis ferredoxin-NADP+ oxidoreductase is capable of functioning as ferric reductase and of driving the fenton reaction in the presence of free flavin. Biometals. 2011;24(1):311-321.
- Hertzberger R, Arents J, Dekker HL, Pridmore RD, Gysler C, Kleerebezem M, et al. H2O2 production in species of the Lactobacillus acidophilus group: a central role for a novel NADH-dependent flavin reductase. Appl Environ Microbiol. 2014;80(7):2229-2239.
- Eschenbrenner M, Coves J, Fontecave M. Ferric reductases in Escherichia coli: the contribution of the haemoglobinlike protein. Biochem Biophys Res Commun. 1994;198(1):127-131.
- Halle F, Meyer JM. Ferrisiderophore reductases of Pseudomonas. Purification, properties and cellular location of the Pseudomonas aeruginosa ferripyoverdine reductase. Eur J Biochem. 1992;209(2):613-620.
- Halle F, Meyer JM. Iron release from ferrisiderophores. A multi-step mechanism involving a NADH/FMN oxidoreductase and a chemical reduction by FMNH2. Eur J Biochem. 1992;209(2):621-627.
- Huyer M, Page WJ. Ferric reductase activity in Azotobacter vinelandii and its inhibition by Zn2+. J Bacteriol. 1989;171(7):4031-4037.
- Kimata S, Mochizuki D, Satoh J, Kitano, K, Kanesaki Y, Takeda K, et al. Intracellular free flavin and its associated enzymes participate in oxygen and iron metabolism in Amphibacillus xylanus lacking a respiratory chain. FEBS Open Bio. 2018;8(6):947-961.
- Le Faou AE, Morse SA. Characterization of a soluble ferric reductase from Neisseria gonorrhoeae. Biol Met. 1994;4(1)126-131.
- Mazoch J, Tesarik R, Sedlacek V, Kucera I, Turanek J. Isolation and biochemical characterization of two soluble iron(III) reductases from Paracoccus denitrificans. Eur J Biochem. 2004;271(3):553-562.
- Moody MD, Dailey HA. Ferric iron reductase of Rhodopseudomonas sphaeroides. J Bacteriol. 1985;163(3):1120-1125.
- Vadas A, Monbouquette HG, Johnson E, Schroder I. Identification and characterization of a novel ferric reductase from the hyperthermophilic Archaeon Archaeoglobus fulgidus. J Biol Chem. 1999;274(51):36715-36721.
- Filisetti L, Valton J, Fontecave M, Nivière V. The flavin reductase ActVB from Streptomyces coelicolor: characterization of the electron transferase activity of the flavoprotein form. FEBS Lett. 2005;579(13):2817-2820.
- Yeom J, Jeon CO, Madsen EL, Park W. Ferredoxin- NADP+ reductase from Pseudomonas putida functions as a ferric reductase. J Bacteriol. 2009;191(5):1472–1479.
- Niederer W. Ferritin: iron incorporation and iron release. Experientia. 1970;26(1):218-220.
- Satoh J, Kimata S, Nakamoto S, Ishii T, Tanaka E, Yumoto S, et al. Free flavins accelerate release of ferrous iron from iron storage proteins by both free flavin-dependent and -independent ferric reductases in Escherichia coli. J Gen Appl Microbiol. 2020;65(1):308-315.
- Weeratunga SK, Gee CE, Lovell S, Zeng Y, Woodin CL, Rivera M, et al. Binding of Pseudomonas aeruginosa apobacterioferritin-ssociated ferredoxin to bacterioferritin B promotes heme mediation of electron delivery and mobilization of core mineral iron. Biochemistry. 2009;48(31):7420-7431.
- Yao H, Jepkorir G, Lovell S, Nama PV, Weeratunga S, Battaile KP, et al. Two distinct ferritin-like molecules in Pseudomonas aeruginosa: the product of the bfrA gene is a bacterial ferritin (FtnA) and not a bacterioferritin (Bfr). Biochemistry. 2011;50(23):5236-5248.
- Massey V. Activation of molecular oxygen by flavins and flavoproteins. J Biol Chem. 1994;269(36):22459-22462.
- Maruyama T, Higuchi K, Yoshida M, Tadano T. Comparison of iron availability in leaves of barley and rice. Soil Sci Plant Nutr. 2005;51(7):1035-1042.
- Nimura Y, Yanagida F, Uchimura T, Ohara N, Suzuki K, Kozaki M, et al. A new facultative anaerobic xylan-using alkalophile lacking cytochrome, quinone, and catalase. Agric Biol Chem. 1987;51(8):2271-2275.
- Niimura Y, Koh E, Yanagida F, Suzuki K, Komagata K, Kozaki M, et al. Amphibacillus xylanus gen. nov., sp. nov., a facultatively anaerobic sporeforming xylan-digesting bacterium which lacks cytochrome, quinone and catalase. Int J Syst Bacteriol. 1990;40(3):297-301.
- Niimura Y, Suzuki K. Bergey’s Manual of Systematic Bacteriology. (2nd edn) Springer, New York, USA. 2009.
- Ohnishi K, Niimura Y, Yokoyama K, Hidaka M, Masaki H, Uchimura T, et al. Purification and analysis of a flavoprotein functional as NADH oxidase from Amphibacillus xylanus overexpressed in Escherichia coli. J Biol Chem. 1994;269(50):31418-31423.
- Mocizuki D, Arai T, Asano M, sasakura N, Watanabe T, Shiwa Y, et al. Adaptive response of Amphibacillus xylanus to normal aerobic and forced oxidative stress conditions. Microbiology. 2014;160(2):340-352.
- Rajagopalan PTR, Grimme S, Pei D. Characterization of cobalt (II)-substituted peptide deformylase: function of the metal ion and the catalytic residue Glu-133. Biochemistry. 2000;39(4):779-790.
- Watanabe A, Yamaguchi T, Murota K, Ishii N, Terao J, Okada S, et al. Isolation of lactic acid bacteria capable of reducing environmental alkyl and fatty acid hydroperoxides, and the effect of their oral administration on oxidative-stressed nematodes and rats. PLoS One. 2020;15(1):e0215113.
- Niimura Y, Ohnishi K, Yarita Y, Hidaka M, Masaki H, Uchimura T, et al. A flavoprotein functional as NADH oxidase from Amphibacillus xylanus Ep01: purification and characterization of the enzyme and structural analysis of its gene. J Bacteriol. 1993;175(24):7945-7950.
- Niimura Y, Poole LB, Massey V. Amphibacillus xylanus NADH oxidase and Salmonella typhimurium alkyl-hydroperoxide reductase flavoprotein components show extremely high scavenging hydroperoxide reductase 22-kDa protein component. J Biol Chem. 1995;270(43):25645-25650.
- Niimura Y, Nishiyama Y, Saito D, Tsuji H, Hidaka M, Miyaji T, et al. A hydrogen peroxide-forming NADH oxidase that functions as an alkyl hydroperoxide reductase in Amphibacillus xylanus. J Bacteriol. 2000;182(18):5046-5051.
- Tartaglia LA, Storz G, Brodsky MH, Lai A, Ames BN. Alkyl hydroperoxide reductase from Salmonella typhimurium. Sequence and homology to thioredoxin reductase and other flavoprotein disulfide oxidoreductases. J Biol Chem. 1990;265(18):10535-10540.
- Niimura Y, Massey V. Reaction mechanism of Amphibacillus xylanus NADH oxidase/alkyl hydroperoxide reductase flavoprotein. J Biol Chem. 1996;271(48):30459-30464.
- Ogura Y. Catalase activity at high concentration of hydrogen peroxide. Arch Biochem Biophys. 1955;57(2):288-300.
- Little C. Steroid hydroperoxides as substrates for glutathione peroxidase. Biochim Biophys Acta. 1972;284(2):375-381.
- Chaudiere J, Tapple AL. Purification and characterization of selenium–glutathione peroxidase from hamster liver. Arch Biochem Biophys. 1983;226(2):448-457.
- Takahasi K, Avissar N, Whitin J, Cohen H. Purification and characterization of human plasma glutathione peroxidase: a selenoglycoprotein distinct from the known cellular enzyme. Arch Biochem Biophys. 1987;256(2):677-686.
- Flohe L, Loschen G, Günzler WA, Eichele E. Glutathione peroxidase, V. The kinetic mechanism. Hoppe-Seyler’s Z Physiol Chem. 1972;353(1):987-999.
- Björnstedt M, Xue J, Huang W, Åkesson B, Holmgren A. The thioredoxin and glutaredoxin systems are efficient electron donors to human plasma glutathione peroxidase. J Biol Chem. 1994;269(47):29382-29384.
- Arai T, Kimata S, Mochizuki D, Hara K, Zako T, Odaka M, et al. NADH oxidase and alkyl hydroperoxide reductase subunit C (peroxiredoxin) fromAmphibacillus xylanus form an oligomeric assembly. FEBS Open Bio. 2015;5(1):124-131.
- Arisaka F, Niimura Y, Minton AP. Comparison of composition-gradient sedimentation equilibrium and composition-gradient static light scattering as techniques for quantitative characterization of biomolecular interactions: A case study. Anal Biochem. 2019;583(2):113339.
- Mochizuki D, Tanaka N, Ishikawa M, Endo A, Shiwa Y, Fujita N, et al. Evolution and diversification of oxygen metabolism of aerotolerant anaerobes in the order Bacillales and other bacterial taxonomic groups. The Bulletin of BISMiS. 2012;3(1):1-17.
- Seaver LC, Imlay JA. Alkyl hydroperoxide reductase is the primary scavenger of endogenous hydrogen peroxide in Escherichia coli. J Bacteriol. 2001;183(24):7173-7181.
- Imlay JA. Where in the world do bacteria experience oxidative stress?. Environ Microbiol. 2019;21(2):521-530.
Citation: Suzuki T, Kimata S, Satoh J, Takeda K, Mochizuki D, Kitano K, et al. (2020) Free Flavin Participates in Iron and Also Oxygen Metabolism in Bacteria. J Bacteriol Parasitol. 11:379.
Copyright: © 2020 Suzuki T, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.