
Indexed In
- Open J Gate
- Genamics JournalSeek
- Academic Keys
- JournalTOCs
- ResearchBible
- Ulrich's Periodicals Directory
- Access to Global Online Research in Agriculture (AGORA)
- Electronic Journals Library
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- SWB online catalog
- Virtual Library of Biology (vifabio)
- Publons
- MIAR
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Research Article - (2019) Volume 10, Issue 1
Dynamics and Fitness Cost of Genetically Engineered Entrobacter cloacae Expressing Defensin for Paratransgenesis in Phlebotomus papatasi
Rangin Abassi, Maryam Akhlaghi, Mohammad Ali Oshaghi*, Amir Ahmad Akhavan, Mohammad Reza Yaghoobi-Ershadi, Rounak Bakhtiary and Fatemeh MohtaramiReceived: 16-Oct-2018 Published: 26-Feb-2019, DOI: 10.35248/2155-9597.1000349
Abstract
Background: Enterobacter cloacae subsp. dissolvens bacterium is a known commensal of the gut microflora of Phlebotomus papatasi, the main vector for zoonotic cutaneous Leishmaniasis, and nominated for paratransgenesis in sand flies. In this study, we evaluated dynamics and fitness costs of engineered E. cloacae for its potential to serve as a 'Trojan Horse' in P. papatasi.
Methods: The engineered strain of E. cloacae transformed with a constantly active expressed red fluorescent protein plus defensin (EC-DR) plasmid and was fed to sand fly colonies via larval food to larvae. A wild type the bacterium (EC-WT) and intact food were used as controls. Fitness characters as well as dynamics of the EC-DR at various development stages of sand fly larvae were tested by plating homogenized specimens and counting fluorescent expressing colonies on the Tet-BHI agar medium.
Results: Enterobacter cloacaeDR producing red fluorescent protein could be isolated from the larvae gut after 36 days when the bacteria were added once in larval pots. The EC-DR with multiple applications had no negative effect on emergence time of instar II larvae, pupae, and adults but increased slightly mortality rate of P. papatasi larvae. The experiment also confirmed lack or weak trans-stadial transmission of E. cloacae DR in P. papatasi. It has minimal fitness cost on P. papatasifeeding behavior and survival.
Conclusion: Results of this study showed that E. cloacae DR is suitable for paratransgenesis of P. papatasi at only adult stage because it did not transmit transstadially.
Introduction
Leishmaniasis is one of the ten major tropical diseases that the World Health Organization (WHO) has been recommending and supporting research on their various aspects. The Leishmaniases are complex diseases of (sub) tropical regions of the world caused by different Leishmania species of Trypanosomatidae and spread by sand flies. The WHO considers the Leishmaniases to be prominent among the global causes of death by infectious diseases [1]. Clinical manifestations produced by Leishmania comprise the visceral (VL) and cutaneous forms. The cutaneous forms of the disease include the cutaneous (CL), diffuse (DCL), and mucocutaneous (MCL) Leishmaniasis, but infections remain asymptomatic in many cases [1-3]. Leishmaniasis is now endemic in 98 countries, and a total of 350 million people are at risk of developing the disease, including 21 countries in the new world and 79 countries in the ancient world. A total of 12 million people suffer from this disease, and occur between 1 and 2.1 million new cases of Leishmaniasis in the world [4].
In Iran, 80% of reported cases are Zoonotic Cutaneous Leishmaniasis (ZCL) form [5]. ZCL is caused by the parasite L. major and transmitted by the sand fly Phlebotomus papatasi . It is endemic in 17 provinces of 31 provinces in Iran [6].
A wide range of Leishmaniasis control methods have been used In Iran, such as indoor residual spraying with DDT, rodenticide, impregnation of bed nets and curtains with deltamethrin [7]. Application of pesticides in addition to undesirable effects on human health and the environment, has led to resistance to sand fly [5,8]. Other interventions such as the L. major autoclavable vaccine with BCG have not been proactive in protecting the disease [9].
Paratransgenesis is an approach in which symbiotic bacteria, fungi, or viruses of the vector insects are genetically manipulated to deliver effector proteins that block development or transmission of the pathogen. This approach attempts to decrease pathogen transmission without adverse effects on vectors themselves. Further, it is considered as a gene delivery mechanism to the host and indigenous bacterial flora of the host vector [10]. The first example of this technique used in the blood sucking bugs [11]. Also bacterial symbionts of tsetse flies, mosquitoes, American cockroach and sand flies have been identified and in some cases successfully transformed and laboratory evaluated to reduce or eliminate carriage of pathogens by host arthropods [12-23].
The midgut bacterial flora of wild-caught insects could be very dynamic and significant fluctuations can be observed depending on the stage of life, nutrients and the physiological age (31, 44). Population structure of symbiotic bacteria is considerably changed post blood meal and gram-negative bacteria will be dominant and could survive in harsh condition of midgut with digestive enzymes (31). Very few of bacteria were only able to transfer from larvae to adult stages because of harsh condition of histolysis processing at pupal stage. Some bacteria are able to colonize in the malpighian tubules or hymolymph (after escaping from midgut wall) and transstadially pass from larvae to adult and presumably remain for long time in the adult gut or hemolymph [16]. Therefore, such symbiotic or commensal bacteria added to the diet of larva insects to pass to adults for paratransgenesis (60).
Enterobacter cloacaebacterium is a species of gram-negative, facultative anaerobic, rod-shaped bacterium belonging to Gammaproteobacteria and Enterobacteriaceae family. The bacteria species limited the development of Plasmodium berghei and P. falciparum by stimulate the immune system of An. stephensi and increases the expression of immune responses compounds such as serine protease inhibitors (SRPN6) (31). The bacterium was found as the natural microflora of different insects including mosquitoes (Anopheles albimanus , An. stephensi , Culex tarsalis , Psorophora columbiae , Aedes triseriatus , Ae. albopictus , and Ae. aegypti ), cockroaches (Periplaneta americana ), as well as sand fly of Ph. papatasi (57). Gonzalez Ceron et al. (51) reported E. cloacae restricted the P. vivax development in midgut of An. albimanus (61-63). E. cloacaehas been transformed with a constantly active expressed red fluorescent protein plus defensin (EC-DR) plasmid. Defensin is an antimicrobial molecule which can kill parasites. Defensins are small cysteine rich cationic proteins and found in plants, vertebrates and invertebrates. They are active against fungi, bacteria, and many viruses. Dehghan showed that EC-DR inhibited P. berghei development in An. stephensi . Maleki-Ravasan et al. (46) showed that the bacterium could obstruct L. major promastigotes in vitro condition. Due to the ability of E. cloacae to direct and indirect prevention of Plasmodium and Leishmania parasites, this bacterium nominated as a candidate for paratransgenesis approach against the malaria and Leishmaniasis. In this study, we evaluated the fitness costs and dynamics of E. cloacaedissolvens expressing red fluorescent and defensin proteins (DR) in midgut of P. papatasi larvae, pupae, and newly emerged adult stages as well as in larval habitats as delivery system for the sand flies in the laboratory condition. The knowledge of bacterium dynamics and its effects in sand flies is necessary before application of the engineered bacterium in the field.
Materials and Methods
Study area
Sand fly sampling was done in July and August 2016 from rural areas of Habib-Abad in Isfahan province. This province is an endemic focus for ZCL in central part of Iran. The sand fly collected with Aspirator from close places to rodent burrows in the night. Live specimens were transferred to laboratory. Rearing of P. papatasisand flies carried out under laboratory condition in 26 ± 2°C and 80% relative humidity with 14:10(L:D) photoperiod in Insectary of School of Public Health, Tehran University of Medical Sciences following the method of Modi & Tesh. Sand fly feeding was carried out with sucrose and honey and blood fed on BALB/c mice.
The bacteria
Enterobacter cloacae subsp. dissolvens was obtained from P. papatasi in the field of ZCL in Isfahan, central Iran [19]. The manipulated strain of E. cloacae contained plasmid expressing difensin and RFP proteins and a gene resistant to Ampicillin.
Dynamics of Enterobacter cloacae -DR in P. papatasi development stages
In order to assess E. cloacae-DR colonization at larval stage, in two separate groups, 107 CFU bacteria per 10 cm diameter of larval growth medium (pot) was added once to the pot at first-instar larvae for the first group, and every 3.5 days for the second group. The presence of the modified bacteria was monitored in the larval instars L1, L2, L3-L4, pupae, and the newly emerged adults. Enterobacter cloacae -DR bacteria were measured by plating the homogenates on LB agar plates and counting the fluorescent colonies using fluorescent microscopy.
In each sampling a number of larvae, pupae, and adults were selected and kept on in the freezer for 10-15 minutes until they were numbed. Then the specimen surfaces were washed with sterile PBS buffer and then their surface bodies were sterilized by ethanol 70% for 5 minutes. After this the individual specimens were washed with PBS buffer. Finally the total body of specimen was transferred to the micro tubes containing 50 ml sterile PBS buffer and homogenized. Then homogenized solution was cultured in the BHI Agar plate contained ampicillin. The plates were incubated at 37°C for 18-20 hours, and total cell formed unites (CFUs) on each plates were counted. The washing solution also was tested for presence of the bacteria ensuring surface sterilization.
Bacterium transstadial transmission
To test the transstadial transmission of E. cloacae -DR bacteria from larvae to adult stage the early and late pupae of both sexes and newly emerged male and female adults were sampled immediately after leaving the puparium. Presence of E. cloacae-DR was tested as above. A number of newly emerged adults were fed either with 20% sucrose and or blood, and existence of E. cloacae-DR was verified in the fed specimens as previously described above.
Results
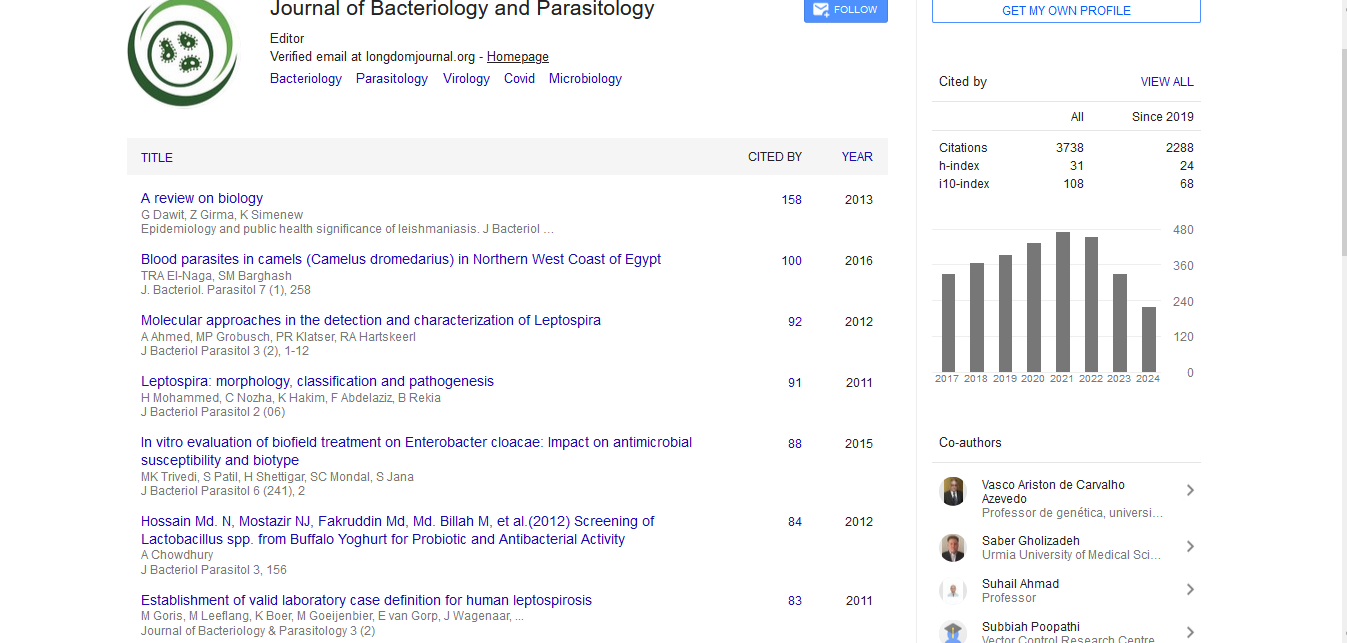
Dynamics of Enterobacter cloacae -DR in larva, pupa, adult P. papatasi
At larval stage, when the engineered bacteria was added to larval pot only one time in the first instar larvae, except for two positive larval specimens with a few colonies, all of the samples including different larval stages (I-IV), pupa and newly emerged adults were negative for E. cloacae D-R. When the bacteria were added every 3.5 days to pots, E. cloacae -RD was found in the guts of P. papatasilarvae (Figures 1 and 2), however, it inadequately transmitted transstadially from larvae to adult stage. On average, the bacterial infection rate in the larval guts and pupae was 24.4% and 22.5% respectively.

Figure 1: Trend of E. cloacae -DR in the gut of Ph. papatasi larvae . 107 CFU bacteria per 10 cm diameter of larval growth medium (pot) were added every 3.5 days.

Figure 2: Average population of E. cloacae –DR in P. papatasi midgut. 107 CFU bacteria per 10 cm diameter of larval growth medium (pot) were added every 3.5 days.
Transstadial transmission
Result of this study showed that the E. cloacae RD was not transmitted transstadial from larvae to adults. Among the adult specimens tested only one post blood-fed specimen harboring 226 CFUs in its gut was positive for the bacterium indicating very low transstadial transmission (Figure 3).

Figure 3: Effect of E. cloacae -DR on Mortality Rate of P. papatasi larvae and pupa. Bars indicate mean and standard error mean (Mean ± SEM). 1 and 3.5 refer to the larvae treated with the bacterium once or every 3.5 days respectively.
Effect of E. cloacae-DR on Mortality Rate of P. papatasi larvae and pupa
At first instar larvae, a greater mortality was observed in the group of sand flies treated with E. cloacae wild-type strain every 3.5 days, although the difference was not statistically significant with other groups. The highest mortality rate was observed in the second to forth instar larvae in all groups. The highest mortality rate was related to the larvae that were treated every 3.5 days with transgenic bacteria and the lowest mortality rate was observed in control group. The differences among three groups were not significant. Also there were no significant differences in the mortality rate of pupae among the three groups (Figure 4).
Figure 4: Effect of E. cloacae -DR on Mortality Rate of P. papatasi larvae and pupa. Bars indicate mean and standard error mean (Mean ± SEM). 1 and 3.5 refer to the larvae treated with the bacterium once or every 3.5 days respectively.
Dynamic trend of E. cloacae population in pot
In this experiment, the bacterial was studied in the experimental medium, which was added to the larval culture media once or every 3.5 days. The observed results showed that the bacterium was present in the paste environment and the results of culture were always positive when bacteria added every 3.5 days. The number of bacteria was often uncountable, and counting the number of colonies was done diluted, and then the number of colonies was counted. In the case of a patch that was added only once at instar-1 larvae, it was observed that over time the number of colonies in the cultured samples decreased and finally reached close to zero after 35 days (Figure 5).

Figure 5: Trend of E. cloacae –DR in the larval pot of P. papatasi.
Discussion
In this study, we first studied the dynamics of Enterobacter cloaca bacteria, a subspecies of dissolvens capable of producing defensin molecules along with a red fluorescent protein (RFP) marker in the P. paparasi sand flies, an important vector of rural Leishmaniasis in Iran and many endemic areas illness in the old world. This bacterium is the candidate of the paratransgenic method to control rural cutaneous Leishmaniasis as well as control of malaria [19,20].
Reviews of Maleki-Ravasan et al. Showed that the microbioum of sand flies Isfahan foci is very rich and there are 40 bacteria species in the gastrointestinal tract of the immature and adult P. papatasi. The researchers were found two species of Bacillus subtilis and Enterobacter cloaca dissolvens subspecies with a high relative abundance in the digestive tube, larval habitats (nest material), and adult food sources. Due to the high correlation of these two bacteria with/by sand flies and living and nonliving factors affecting the epidemiology of rural cutaneous Leishmaniasis and the natural rotation/circulation of these two bacteria between sand fly vectors, the disease reservoir, the Salsola plant, supplies sugar sources for vectors and food for the reservoirs, and the rodent feces supplying/providing larval food vectors, were introduced these two bacteria as two suitable paratransgenic candidate sand flies. Due to the failure of the transgenic bacteria B. subtilis in the study, focused on bacteria Enterobacter cloacae[19]. The bacteria were successfully modified/manipulated and the genes producing the two proteins, defensin and red fluorescence/ fluorescent, were added to the Bacteria genome. Enterobacter cloacae are natural human and animal digestive bacteria and are not considered as a primary pathogen [24]. This Bacterium is found in water, sewage and soil [25]. This bacterium is the most abundant bacteria in the gut of insects of medical importance [26]. These bacteria are found in the gut of insects such as Aedes albopictus and Ae. aegypti , An. gambiae , An. albimanus and in P. papatasi sand flies [19,27]. This Bacteria is an optional anaerobic and therefore can be adapted to with different environments [25]. This Bacterium produces hemolysin against some of the parasites [28]. It is also a rhizobacter and stimulates plant growth [29]. The present study investigates the effect and determination of population dynamics ofEnterobacter cloacae in the on immature stages of P. papatasi
In this study, Enterobacter cloacae bacteria produce defensin molecules. This molecule is a plant defensin and able to kill the parasites. Defensin of insects and mammals, unlike plant defensins are active against bacteria. Plant defensin often act against fungi. They are not only active against pathogenic fungi of plants (such as Fusarium culmorum and Botrytis cinerea ) but also are against human pathogenic yeast and fungi (such as C. albicans ). Plant defensins inhibit trypsin and alpha-amylase enzymes. Trypsin enzymes are involved in blood digestion swallowed and the survival of the parasite in the early stages of the conversion of amastigotes to promastigotes. Alpha amylase plays a role in the digestion of carbohydrates in plants sap.
Defensin used in this study (RsAFP1), which is related to radish plant, is classified in the ninth and third subgroup of plant defensin. Defensin protein (Rs-AFPs 0.5%) forms the whole protein of radish seeds. During germination, the seed loses its protective coating and is exposed to soil microorganisms. Rs-AFPs are released after fusing the protective coating and make up 30% of the protein released from the seed coating. This amount of released proteins is such that they suppress the growth of fungi in the soil.
The study found that Enterobacter cloacaeis not stable in the liver containing diet and is rapidly disappearing. According to previous studies, liver has been found to contain vitamin C. This vitamin has anti-bacteria properties. In a study of 148 animals, it was found that vitamin C can reduce the contamination of bacteria, viruses and protozoa, or completely prevent inflammation [30]. That's why it's likely that liver or its composition with larval food produces metabolites prevent the growth of the bacteria in the food. The results of our study showed that the number of bacteria increased when the liver was removed from the food, and the bacteria stability was much better.
In a survey of attractiveness of larval food containing the bacteria, we also observed that larvae do not absorb bacteria food; on the other hand, bacteria do not have a high stability in the pot and rapidly decrease their population, which may be due to environmental humidity/ambient humidity or other causes. These two factors primarily affects in the receiving bacteria in gut of larvae, at a later stage if the bacteria enters the gut larvae, the number of bacteria inside the larvae is greatly reduced, it is may be due to the unfavorable conditions of the gut, pH, presence of various enzymes, immune system of insects, lack of appropriate food conditions or the presence of other microorganisms.
The results of this study showed that E. cloacaebacteria are not stable in Pot (growth environment of larvae of sand flies) and over time; their populations are reduced, which may be due to various factors, including moisture/humidity in the pot environment. It seems that high humidity prevents the growth of these bacteria which was also observed in the other study [20]. They showed that the population of E. cloacae bacteria in Anopheles larva habitat (water) drastically reduced. Reduce the population of bacteria in a pot indirectly reducing the number of bacteria in the gut of the sandflies larvae. In addition, the lack of attractiveness of bacteria for larvae also reduces the amount of bacteria in the larvae. In the test of attractiveness of the food containing the bacteria E. cloacaewas found that P. papatasisand flies larvae were not attracted into the bacteria contaminated food. The repellent/repulsion/defatting property can be due to the undesirable odor of the bacteria and the fragrance of the larval and bacteria food complex. The results of this study showed that E. cloacae bacteria did not increase significantly in the gut of larvae.
The lack of attractiveness of the bacteria for larvae and the bacteria instability in the pot due to external factors, and the adverse/ unfavorable physical and chemical conditions of the larvae gut, pH, the presence of various enzymes, insect immunity, lack of suitable food sources for Bacteria growth, the presence of other microorganisms are internal factors Influencing the E. cloacae bacteria is inside the larvae.
In this study, we used two different treatments to infect larvae and infective larval food feed larvae from bacteria contaminated food. In the first treatment only once and in the second treatment, every 3.5 days, the infected food was added to the pot. The results showed that the bacteria were not adapted to the laboratory conditions in the larval growth medium. In the first treatment, the bacteria gradually decreased, and on day 36 it was close to zero. But in the second treatment, there was a slight decrease in the number of bacteria over time, but the number of bacteria, even after 50 days, never dropped below 10,000/did not fall below ten thousand. So if these bacteria are used to infect larvae in a paratransgenic, infecting the larva environment should be repeated at least once a week, which is not very appropriate and may not be operational in the desert.
In this study, it was found that as the age of larvae increases, the amount of bacteria increases in the larvae gut. And the maximum number of bacteria was observed at the end of age 3 and early larval age 4. And as the age of larva increases, the number of bacteria gradually decreases as it approaches the pupa. Maximum nutrition is done in larvae of age 3 and early age 4. And the amount of nutrition has a direct impact on the bacteria eaten and inserted into the insect gut there is a direct relationship between the age and feeding activity of the larvae with the number of bacteria found in their gut.
In investigate the effect of transgenic and wild strains of E. cloacae the growth rate of larvae age 2, with two different diets/regimes, adding one or more than one bacteria into the pot, it was found thatE. cloacaebacteria has no effect on the time of the emergence of the second instar larvae. This point, with the advice of researchers, seems appropriate to believe that the candidate bacteria for paratransgenic should not have a negative effect on insect adaptation but the effect of bacteria on the emergence of pupae and adult, it was found that the Bacteria strain of transgenic and wild E. cloacae delayed the emergence of pupae and adult diets is once bacteria adding. In justifying this phenomenon, the effect of bacteria inhibition on larval nutrition can be mentioned. It seems that food containing bacteria has a negative effect on larval nutrition therefore, feeding the larvae was inappropriate and consuming a small amount of food at each feeding time, which is why their growth and development period has been prolonged. When larvae were fed with bacteria every 3.5 days, it was observed that the transgenic E. cloacae diet had no effect on the appearance of pups, but the wild strain of the bacteria prolonged the time of emergence of pupae and mature. It seems transgenic bacteria producing antimicrobial defensin is that other bacteria in the environment, destroys or reduces their population. Some of these bacteria are harmful to the growth of larvae. Therefore, the presence of transgenic bacteria and the production of defensin reduce bacteria and even fungi, and thus the growth trend in these insects does not differ significantly from the control group but the wild strain of bacteria that does not produce any defensin and the inhibitory bacteria of growth remain intact, and overall the larvae grow longer. In our study, based on the observations, it seems that the delay occurred to convert larvae to pupae in the pre-pupa stage. Studies show that the minimum number of bacteria in the pot environment is more than 10,000 per square millimeter of the pot. Possibly, the olfactory receptors of the insect are saturated and the masking happens and the insect does not react to the smells and bacteria and behaves like the control group. The wild strain Bacteria does not have defensin, which is why the bacteria agent itself has a negative effect, causing a delay in growth, and the effect of defensin in saturated state can be considered to justify this phenomenon. There may, of course, be other reasons to justify this situation, which requires more investigation.
In this study, it was found that when E. cloacae transgenic bacteria were added to the peripheral environment every 3-5 days, the mortality rate increased in larvae. Mortality also occurs in larvae of age 2-4. It seems that transgenic bacteria, producing defensin, or affecting the level of larval nutrition, have an adverse effect on larval consistency and survival. However, E. cloacae bacteria did not show a significant difference in the mortality rate with the control group. In the stage of pupal, the mortality rate between different groups is not statistically different. The main reason for this is the lack of transstadial and the absence of bacteria in the pupal stage, and different treatments are similar in the presence or absence of bacteria condition, and our studies showed that in most groups, most puppies are bacteriologically negative. Therefore, the There is no difference between the control and transgenic groups and the wild strain [22-31].
Conclusions
In this study, it was found that E. cloacae bacteria are not transstadial from larvae to mature P. papatasi , or transmission of larvae to mature animals is negligible. In this study it was found that only one of the 70 adults tested was positive. The lack of transstadial of E. cloacae bacteria is an undesirable point for using these bacteria in the paratransgenic strategy against P. papatasisandflies. In practice, these bacteria cannot be used in immature stage of sand flies, and only can be used in adult stage. In the study of Dehghan et al. on Anopheles stephesi bacteria, it was observed that the bacteria in the Anopheles was not transstadial or very weak. The lack of transstadial of bacteria according to the results of other studies, which all acknowledge that most bacteria in the pupa stage under the influence of enzymes and adverse conditions (histolysis) lose their survival and very few bacteria that, can be transstadial. Most transstadial bacteria are of the intracellular type, such as: Asaia bacteria in mosquitoes An. Stephensi and Wolbachia bacteria. However, some extracellular bacteria such as Seratia in Anopheles stephensi and B. subtilus bacteria in P. argentipes sand flies, Pseudomonas bacteria also occur in An. stephensi are transstadial.
Results of this study showed that due to the incompatibility of E. cloacae bacteria in a pot with a single use, lack of sufficient growth in larvae and its lack of transstadial, E. cloacae is not suitable for paratransgenesis of P. papatasiat larval stage and should be used at adult stage.
REFERENCES
- http://www.who.int/leishmaniasis/en/
- Ostyn B, Gidwani K, Khanal B, Picado A, Chappuis F, et al. (2011) Incidence of symptomatic and asymptomatic Leishmania donovani infections in high-endemic foci in India and Nepal: A prospective study. PLoS Negl Trop Dis 5: 1284.
- Andrade-Narvaez FJ, Loría-Cervera EN, Sosa-Bibiano EI, Van Wynsberghe NR (2016) Asymptomatic infection with American cutaneous leishmaniasis: epidemiological and immunological studies. Mem Inst Oswaldo Cruz 111: 599–604.
- World Health Organization (2010) Control of the Leishmaniasis: Report of A Meeting of the WHO Expert Committee on the Control of Leishmaniases. Geneva, Switzerland.
- Shirani-Bidabadi L, Zahraei-Ramazani A, Yaghoobi-Ershadi MR, Rassi Y, Akhavan AA, et al. (2017) Assessing the insecticide susceptibility status of field population of Phlebotomus papatasi (Diptera: Psychodidae) in a hyperendemic area of zoonotic cutaneous leishmaniasis in Esfahan Province, Central Iran. Acta Trop 176: 316-322.
- Yaghoobi-Ershadi M (2012) Phlebotomine Sand Flies (Diptera: Psychodidae) in Iran and their role on Leishmania transmission. J Arthropod Borne Dis 6: 1-17.
- Yaghoobi-Ershadi MR (2016) Control of phlebotomine sand flies in Iran: A review article. J Arthropod Borne Dis 10: 429-444.
- Seyedi-Rashti MA, Nadim A (1974) Attempts to control zoonotic cutaneous leishmaniasis in the Isfahan area, Iran. Iran J Publ Hlth 2: 199–203.
- Momeni AZ, Jalayer T, Emamjomeh M, Khamesipour A, Zicker F, et al. (1999) A randomised, double-blind, controlled trial of a killed L. major vaccine plus BCG against zoonotic cutaneous leishmaniasis in Iran. Vaccine 17: 466-472.
- Dillon RJ, Dillon VM (2004) The gut bacteria of insects: nonpathogenic interactions. Annu Rev Entomol 49: 71–92.
- Durvasula RV, Gumbs A, Panackal A, Kruglov O, Aksoy S, et al. (1997) Prevention of insect-borne disease: An approach using transgenic symbiotic bacteria. Proc Natl Acad Sci U S A 94: 3274–3278.
- Medlock J, Atkins KE, Thomas DN, Aksoy S, Galvani AP (2013) Evaluating paratransgenesis as a potential control strategy for African trypanosomiasis. PLoS Negl Trop Dis 7: 2374.
- Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, et al. (2007) Bacteria of the genus asaia stably associate with anopheles stephensi, an asian malarial mosquito vector. Proc Natl Acad Sci USA 104: 9047–9051.
- Chavshin AR, Oshaghi MA, Vatandoost H, Pourmand MR, Raeisi A, et al. (2012) Identification of bacterial microflora in the midgut of the larvae and adult of wild caught AnoPeles stePensi: a step toward finding suitable paratransgenesis candidates. Acta Trop 121: 129–34.
- Chavshin AR, Oshaghi MA, Vatandoost H, Pourmand MR, Raeisi A, et al. (2014) Isolation and identification of culturable bacteria from wild anopheles culicifacies, a first step in a paratransgenesis approach. Parasit Vectors 7: 419.
- Wang S, Dos-Santos ALA, Huang W, Liu KC, Oshaghi MA, et al. (2017) Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science 357: 1399-1402.
- Akbari S, Oshaghi MA, Hashemi-Aghdam SS, Hajikhani S, Oshaghi G, et al. (2015) Aerobic bacterial community of american cockroach periplaneta americana, a step toward finding suitable paratransgenesis candidates. J Arthropod-Borne Dis 9: 35–48.
- Hillesland H, Read A, Subhadra B, Hurwitz I, McKelvey R, et al. (2008) Identification of aerobic gut bacteria from the kala azar vector, phlebotomus argentipes: a platform for potential paratransgenic manipulation of sand flies. Am J Trop Med Hyg 79: 881–886.
- Maleki-Ravasan N, Oshaghi MA, Afshar D, Arandian MH, Hajikhani S, et al. (2015) Aerobic bacterial flora of biotic and abiotic compartments of a hyperendemic Zoonotic Cutaneous Leishmaniasis (ZCL) focus. Parasit Vectors 8: 63.
- Dehghan H, Oshaghi MA, Moosa-Kazemi SH, Yakhchali B, Vatandoost H, et al. (2017) Dynamics of transgenic enterobacter cloacae expressing green fluorescent protein-defensin (GFP-D) in anopheles stephensi under laboratory conditions. J Arthropod Borne Dis 11: 515-532.
- Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, et al. (2012) Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc Natl Acad Sci U S A 109: 12734-9.
- Capone A, Ricci I, Damiani C, Mosca M, Rossi P, et al. (2013) Interactions between Asaia, Plasmodium and Anopheles: new insights into mosquito symbiosis and implications in malaria symbiotic control. Parasit Vectors 6: 182.
- Hurwitz I, Hillesland H, Fieck A, Das P, Durvasula R, et al. (2011) The paratransgenic sand fly: A platform for control of Leishmania transmission. Parasit Vectors 4: 82.
- Keller R, Pedroso MZ, Ritchmann R, Silva RM (1998) Occurrence of virulence associated properties in Enterobacter cloacae. Infect Immun 66: 645-649.
- Nishijima, K A, Sanders Jr WE, Sanders, CC (1997) Entrobacter spp: Pathogens poised to flourish at the turn of the century. Clin Micro Boil Rev 10: 220-241.
- Gouveia C, Asensi MD, Zahner V, Range EF, Oliveira SM (2008) Study on the bacterial midgut microbiota associated to different Brazilian populations of Lutzomyia longipalpis (Lutz & Neiva) (Diptera: Psychodidae). Neotrop Entomol 37: 597-601.
- Cirimotich CM, Ramirez JL, Dimopoulos G (2011) Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe 10: 307-310.
- Hertle R, Hilger M, Weingardt-Kocher S, Walev I (1999) Cytotoxic action of Serratia marcescens hemolysin on human epithelial cells. Infect Immun 67: 817-25.
- Liu WY, Wong CF, Chung KMK, Jiang JW, Leung FCC (2013) Comparativvvve genome analysis of Enterobacter cloacae. PLOS One 8: 74487.
- Hemilä H (2017) Vitamin C and Infections. Nutrients 9: 339.
- Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, et al. (2009) A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 139: 1268–1278.
Citation: Abassi R, Akhlaghi M, Oshaghi MA, Akhavan AA, Yaghoobi-Ershadi MR et al. (2018) Dynamics and Fitness Cost of Genetically Engineered Entrobacter cloacae Expressing Defensin for Paratransgenesis in Phlebotomus papatasi. J Bacteriol Parasitol 10:349. doi: 10.4172/2155-9597.10.349
Copyright: © 2019 Abassi R, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.